
Nocturnal transpiration of Populus euphratica authenticated by measurements of stem sap flux, leaf gas exchange and stomatal microsturcture
-
摘要: 不完全的气孔关闭引起的夜间蒸腾在不同物种和环境中普遍存在,且其大小与水汽压差和土壤水分有效性正相关,这意味着荒漠河岸林是研究夜间蒸腾的理想区域。本文基于木质部液流、叶片气体交换、显微结构及环境因子测定证实了胡杨夜间蒸腾的存在:1)夜间叶片气孔是不完全关闭的,平均气孔导度为45 mmol/(m2·s),远大于文献报道的杨属最小气孔导度(约为5 mmol/(m2·s)),平均蒸腾速率为0.7 mmol/(m2·s),两者分别占白天的26%和17%,这表明高的气孔导度和蒸腾速率主要是气孔开放引起;2)木质部平均液流速率白天为31.3 cm/h,夜间为16.5 cm/h,约为白天的53%,无论是白天还是夜间,液流速率与水汽压差均呈显著的对数关系,水汽压差可以解释55%的夜间液流变化,这表明夜间液流由蒸腾和组织补水两部分组成,因此,如何将夜间液流区分为夜间蒸腾和组织补水还有待进一步研究。午夜后液流速率的增加与木质部水势和径向生长变化是一致的,而与水汽压差是相反的,说明午夜后液流速率的增加是组织补水而非蒸腾。Abstract: Nocturnal transpiration, as a consequence of incomplete stomatal closure, is prevalent across species and environments, and high nocturnal atmospheric vapor pressure deficit (VPD) along with high soil water availability are the most commonly reported environmental drivers of it, which is coincided with the conditions of riparian forest in extremely arid region. Based on the measurements of xylem sap flux, leaf gas exchange, stomatal microstructure and environmental factors, we confirmed the occurrence of nocturnal transpiration in P. euphratica from followings: 1) incomplete stomatal closure was observed and mean stomatal conductance was 45 mmol/(m2·s), greater than the minimum stomatal conductance of Populus spp. reported in the literature (approximately 5 mmol/(m2·s)), and along with mean transpiration of 0.7 mmol/(m2·s), both accounting for 26% and 17% of daytime, respectively. This suggested that high stomatal conductance and transpiration are largely resulted from stomatal opening. 2) On average, sap velocity was 31.3 cm/hour at daytime and 16.5 cm/hour at nighttime, which accounting for 53% of daytime. Whether during daytime or nighttime, sap velocity was logarithmic positively related with VPD, and this could explain 55% of nighttime sap velocity change, suggesting that nighttime sap flow was composed of transpiration and tissue refilling. Thus, further research to distinguish the nocturnal transpiration and tissue refilling is needed. Noteworthly, the abruptly increased sap flow after midnight was synchronized with the stem radius change and water potential, but oppositely with VPD, showing that increased sap flow after midnight was induced by tissue refilling but not transpiration.
-
传统植物学气孔调节最优化理论认为:夜间,大部分的C3和C4植物都会因无法通过光合作用获取CO2而关闭气孔,因此在夜间不会发生蒸腾作用[1]。但是,大量观测表明一些植物夜间气孔并不是完全关闭的[2]。夜间蒸腾在各种物种和环境中是普遍存在的,且其主要与水汽压差和土壤水分有效性呈显著正相关[2-5]。这意味着生长于气候干旱(高水汽压差)、水分充足(高土壤水分有效性)条件下的荒漠河岸植物可能存在夜间蒸腾。已有研究表明,广泛生长于美国西南部干旱、半干旱区河岸的中国柽柳(Tamarix chinensis)[6]、窄叶杨(Populus angustifolia)和大叶钻天杨(Populus balsamifera)均存在夜间蒸腾[7],且杨属(Populus spp.)植物在干旱条件下能维持高的夜间气孔导度[8]。那么,在干旱区内陆河流域以胡杨(Populus euphratica)和柽柳(Tamarix spp.)为建群种的荒漠河岸林是否也存在类似的夜间蒸腾现象, 笔者采用涡度协方差技术证实柽柳存在夜间蒸腾[9],但对于胡杨由于缺乏有关方面的深入研究,目前尚无定论。
目前植物夜间蒸腾研究主要集中在叶片尺度、单株尺度和冠层尺度[10]。首先,气孔是植物叶片与大气环境进行气体交换的通道,如果夜间蒸腾发生,则可通过直接观察气孔是否开放即叶片显微结构观察判断夜间蒸腾发生的可能,同时也可通过气体交换参数(如气孔导度和蒸腾速率)测定以确定夜间蒸腾是否存在,因此该方法最早在确定夜间蒸腾中得到广泛应用[1, 5-6]。其次,如果夜间蒸腾发生必然引起木质部液流的变化,因此可通过测定木质部液流速率判断夜间蒸腾是否发生。近年来,迅速发展的热比率法(HRM)和热场变形法(HFD)等由于能测定低速和负向液流而在夜间蒸腾研究中得到广泛应用[4, 11-14]。因此,本研究以黑河下游荒漠河岸建群种胡杨为研究对象,拟从木质部液流、叶片气体交换参数测定和气孔显微结构观测等多手段证实胡杨夜间蒸腾的存在。
1. 研究区概况
研究区位于黑河下游额济纳胡杨林自然保护区。该区气候极端干旱[15],据额济纳气象站1961—2012年资料(图 1):气温呈增加趋势,平均年增长率为0.046 ℃/a,最高气温(白天)年增长率最小,为0.026 ℃/a,最低气温(夜间)年增长率最大,为0.070 ℃/a,这与全球气候变化趋势是一致的[16];而相对湿度呈降低趋势,平均年降低率为0.065%/a。因此,研究区水汽压差应该是增加的,尤其是夜间,这极可能引起夜间蒸腾的发生。
![]() 图 1 额济纳气象站1961—2012年日平均(Mean)、最高(Max.)、最低(Min.)气温(Ta)及相对湿度(RH)变化Figure 1. Changes of daily mean, maximum, minimum air temperature (Ta) and relative humidity (RH) in 1961-2012 from Ejin Meteorological Station
图 1 额济纳气象站1961—2012年日平均(Mean)、最高(Max.)、最低(Min.)气温(Ta)及相对湿度(RH)变化Figure 1. Changes of daily mean, maximum, minimum air temperature (Ta) and relative humidity (RH) in 1961-2012 from Ejin Meteorological Station2. 材料与方法
2.1 试验材料
本试验依托中国科学院旱区寒区环境与工程研究所阿拉善荒漠生态水文试验研究站(以下简称“阿拉善站”),地理坐标为42°02′N、101°02′59″E,海拔925 m。试验场设在距离研究站西南方向200 m处,面积为100 m×100 m,试验场内优势种为胡杨,林下冠层内零星分布有灌木多枝柽柳(Tamarix ramosissima)和草本苦豆子(Sophora alopecuroides)、花花柴(Karelinia caspia)等。试验场内胡杨群落特征描述详见文献[17]。
2.2 树干液流速率及环境因子测定
于2012年4—10月,选取人为影响小、个体大小基本一致,冠形匀称,无偏冠及严重枯枝现象的胡杨标准木2株作为试验对象,采用热比率法(Heat Ratio Method)测定木质部液流速率(Vs,cm/h),安装位置为树干木质部北向胸径1.3 m处,液流速率计算及位置偏移和伤口校正具体参见文献[15, 17]。同时,在试验区内安装自动气象站一套用于环境因子连续观测,主要观测指标包括:光量子通量密度(PAR,mmol/(m2·s),QS2,ICT,AU)、空气温湿度(Ta,℃;RH,%;083D-1-35,USA)等。所有观测项目的传感器以电缆同数据采集器相连,自动数据采集器(Zeno3200-A-D,USA)对以上项目每30 min记录1次。水汽压差(VPD,kPa)计算根据Ta和RH [18],白天和夜间划分根据PAR大于还是等于0 mmol/(m2·s)。由于胡杨为国家二级保护物种,其“真实零值(Ture Zero)”取夜间水汽压差接近于0 kPa时的液流速率,约为4cm/h(图 2)。
![]() 图 2 胡杨树干液流速率(Vs)及水汽压差(VPD)日变化2012年7月25—28日,图中阴影部分表示夜间。Figure 2. Daily variation of sap flow velocity (Vs) of Populus euphratica and VPDThe shaded area represent the night-time (25-28, July 2012).
图 2 胡杨树干液流速率(Vs)及水汽压差(VPD)日变化2012年7月25—28日,图中阴影部分表示夜间。Figure 2. Daily variation of sap flow velocity (Vs) of Populus euphratica and VPDThe shaded area represent the night-time (25-28, July 2012).于2015年6—9月在阿拉善站内选择4棵幼树(树龄为8年)采用直径测定仪(DD-L, Ecomatik, Munich, Germany)测定径向生长变化(Rd, μm),并于其中1棵树地面以上75cm高度处安装树干液流仪测定液流速率Vs,同时在液流仪上、下各50cm处安装茎干水势仪(PSY1, ICT International Pty Ltd., Australia)测定其木质部水势(Ψs, MPa)[19],测定频率为30 min。PSY1水势仪安装前,先用小刀将拟安装部位削平,以平面与水势仪探头之间无间隙为佳,然后用去离子水将削去表皮的茎干表面冲洗3~5次,再用可吸水滤纸搽拭干净,之后在水势仪外圈抹上凡士林后安装于经处理的茎干部位,并用特定支架固定后用锡箔纸整个包裹。PSY1茎干水势仪包括2个串联式焊接镍铬-康铜热电偶,它们位于一个有镀铬外层的黄铜腔内,形成一个大的热隔离块。在黄铜腔内,一个热电偶与茎干样本相接触,另一个热电偶同时测量黄铜腔内的空气温度,然后在珀耳帖冷却脉冲之后测量湿球温差。另外,还有一个焊接铜-康铜热电偶位于黄铜腔内用于测量仪器温度从而实现温度补偿。基于水势与温度梯度关系,所有测定的温度用于计算植物茎干水势(Ψ):
ψ=(WBD/(C1Tc+C2)−CI)/−CS+ΔT/k⋅CF (1) 式中:WBD为湿球温度(℃);Tc为黄铜腔内温度(℃);CI为校准曲线截距;CS为校准曲线斜率;C1和C2为温度校准经验常数;ΔT为腔内热电偶和样品热电偶温度差(℃);k为镍铬热电偶输出;CF为ΔT的校准因子。在安装前,每个PSY1采用不同浓度(0.1、0.2、0.3、0.4、0.5、1.0mol/L)NaCl标准液单独校准,得到CI和CS。
2.3 叶片气体交换测量
于2014年7—9月在阿拉善站胡杨试验地选择幼树4株作为观测样树,平均高度为3.0 m,地面0.5 m平均胸径为48.4cm,平均冠幅为2.1 m×2.1 m。每株选取3片不同方位健康、完整叶标记,采用Li-6400XT(LI-COR Biosciences, Nebraska, USA)测定叶片气孔导度(gs,mmol H2O/(m2·s))、蒸腾速率(Tr,mmol H2O/(m2·s))等叶片气体交换参数。测定日期为2014年7月17—18日,7月31日—8月1日和9月2—3日,测定频率为2 h。
2.4 叶片气孔显微结构观察
于2015年5—9月的1—3日选择阿拉善站胡杨试验地内叶片繁茂的幼树3株作为叶片离体采集,每个样树采集叶片3个,置于F.A.A.固定液(38%甲醛5 mL:冰醋酸5 mL:70%酒精90 mL)中固定至完全褪绿,置于液氮罐中冷藏带回。白天从08:00—20:00,每2 h取样1次,夜间从00:00—06:00,每1 h取样1次。所有采集样品在兰州大学生命科学学院试验中心用“透明胶带粘取法”制片[20],后用扫描电镜(OLYMPUS,S-3400N,Japan)观察并拍照,每个叶片随机观察3个视野,白天每个视野取200 μm和100 μm各1次,夜间增加50 μm 1次。
2.5 数据分析
由于受仪器限制,2015年观测的包括叶片气体交换参数、树干液流、树干水势、径向生长和气孔显微结构等,2012年仅观测了树干液流,但时间序列较为完整,2014年测定了不同月份胡杨叶片气体交换参数。2014年和2015年观测均在阿拉善站试验场内,2012年位于阿拉善站旁500 m老胡杨林内,土壤-水文条件基本一致,而气象因子除降雨和相对湿度在0.05显著性水平下存在显著差异外,其他影响蒸腾的要素(如水汽压差、光照和风速)均无显著差异,不同来源数据具有可比性。因此,本文分析数据以2014和2015年为主,2012年观测数据作为补充。
3. 结果与分析
3.1 木质部液流速率
胡杨木质部液流速率和水汽压差连续3 d日变化过程如图 2所示。由图可见,胡杨液流存在明显的昼夜节律性,白天木质部液流从06:00开始迅速增加至正午后14:00达到最大值,之后迅速降低,其变化趋势与水汽压差变化基本一致;夜间,液流速率不为0,在7月26日液流速率从日落至黎明前呈逐渐降低趋势,而7月27日液流在23:00后呈波动上升趋势,这与水汽压差的增加相一致,但夜间01:30后水汽压差开始下降,但液流速率仍在上升。正常条件下,随着夜间气孔部分关闭,液流速率应呈逐渐减小趋势,但由于夜间水汽压差的增加导致液流速率并未如预测的呈减小趋势(图 2中虚线所示)。
表 1 试验期内额济纳站气象要素变化Table 1. Changes in meteorological factors in Ejin Meteorological Station during experiment period年份
Year降水量
Precipitation (P)/mm日照时间
Sunshine duration(N)/h气温
Air temperature(Ta)/℃相对湿度
Relative humidity(RH)/%水汽压差
Vapor pressure difference (VPD)/kPa风速
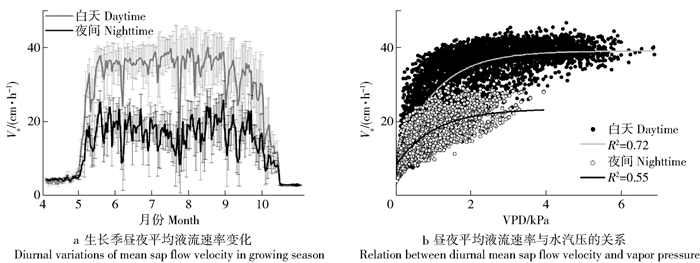
Wind speed(U)/(m·s-1)2012 32.7 8.77±3.04 9.44±14.47 27.72±12.23 1.66±1.27 2.60±1.09 2014 17.2 8.33±3.15 10.55±13.30 30.16±12.14 1.70±1.21 2.63±1.11 2015 44.4 8.63±3.07 10.66±12.78 32.05±14.58 1.71±1.22 2.68±1.11 Sig. No No Yes No No 2012年胡杨白天和夜间液流速率变化如图 3a所示。由图可见,白天和夜间变化趋势一致,展叶期之前(5月4日)和落叶期之后(10月15日)白天和夜间液流速率无显著差异,在此期间,液流速率白天平均为(31.3±9.7)cm/h,夜间平均为(16.5±5.0)cm/h,后者约为前者的53%。液流速率与水汽压差的关系如图 3b所示。由图可见,无论是白天还是夜间,液流速率随着水汽压差的增加呈先增加后稳定的趋势,两者的关系符合对数函数,决定系数(R2)白天大于夜间。但是,无论是白天还是夜间,木质部液流速率与风速无关。
![]() 图 3 胡杨2012年生长季白天和夜间平均液流速率(Vs)变化及其与水汽压差(VPD)的关系Figure 3. Variations of mean Vs of daytime and night-time and the relationship between Vs and VPD during the growing season in 2012 for P. euphratica
图 3 胡杨2012年生长季白天和夜间平均液流速率(Vs)变化及其与水汽压差(VPD)的关系Figure 3. Variations of mean Vs of daytime and night-time and the relationship between Vs and VPD during the growing season in 2012 for P. euphratica3.2 叶片气体交换参数
2014年7—9月典型日胡杨叶片gs和Tr的变化如图 4所示。由图可见,叶片gs呈先增大后减小的趋势,在上午08:00—12:00之间达到最大值,夜间gs不为0,甚至在黎明前呈增大趋势。整体而言,白天gs在45~430 mmol/(m2·s)之间变化,平均为175 mmol/(m2·s);夜间gs在10~160 mmol/(m2·s)之间变化,平均为45 mmol/(m2·s),夜间gs约为白天gs的26%。Tr变化与gs基本一致,但最大值出现在正午或午后14:00左右,夜间Tr同样不为0,平均为0.7 mmol/(m2·s),约为白天Tr的17%。
![]() 图 4 胡杨叶片气孔导度(gs)和蒸腾速率(Tr)典型日变化(2014年7月17—18日,7月31日—8月1日和9月2—3日)Figure 4. Typical daily variations of stomatal conductance (gs) and transpiration rate (Tr) of P. euphratica during July 17 to 18, July 31 to August 1 and September 2 to 3, 2014
图 4 胡杨叶片气孔导度(gs)和蒸腾速率(Tr)典型日变化(2014年7月17—18日,7月31日—8月1日和9月2—3日)Figure 4. Typical daily variations of stomatal conductance (gs) and transpiration rate (Tr) of P. euphratica during July 17 to 18, July 31 to August 1 and September 2 to 3, 20143.2 叶片气孔显微结构
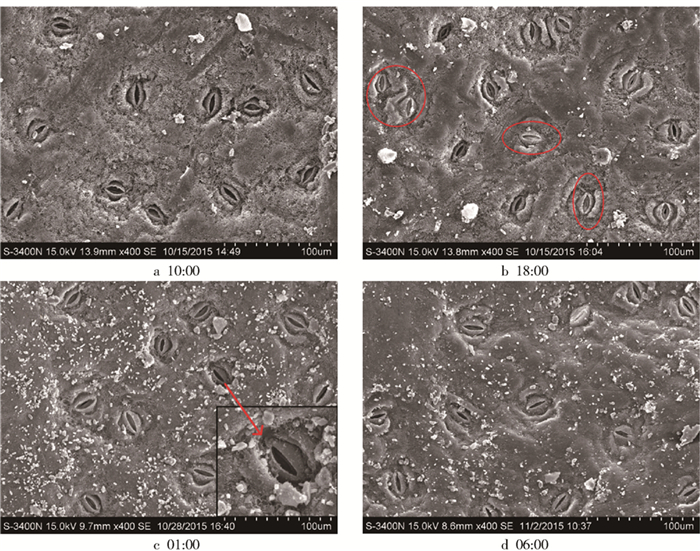
研究表明,夜间蒸腾是由于气孔的不完全关闭引起的,如果夜间发生蒸腾,其气孔必然是开放的且能观测到的。叶片气孔显微结构观察表明(图 5):在光合最强的10:00(a)胡杨叶片气孔基本全部开放,18:00(b中红色区域)出现气孔部分关闭,次日01:00(c)仍有部分开放但气孔开度明显减小,甚至在黎明前06:00(d)开放的气孔略有增加。可见,胡杨叶片的气孔在夜间不完全关闭,这就为水分从气孔散失提供了可能。
![]() 图 5 胡杨幼树白天和夜间叶片气孔显微结构变化(2015年7月1—2日)Figure 5. Changes of leaf stomatal microstructure of P. euphratica at daytime and nighttime during July 1 to 2, 2015
图 5 胡杨幼树白天和夜间叶片气孔显微结构变化(2015年7月1—2日)Figure 5. Changes of leaf stomatal microstructure of P. euphratica at daytime and nighttime during July 1 to 2, 20154. 讨论
4.1 夜间蒸腾存在的证据
气孔是植物叶片与大气环境进行气体交换的通道,夜间气孔是否开放是判断夜间蒸腾存在的前提,但对于夜间不完全气孔关闭究竟是向上的蒸腾作用还是向下的叶片吸水存在争议[21-22]。若是前者,则可通过测定叶片气孔导度和蒸腾速率判断夜间蒸腾是否发生[2-3, 5, 7, 23]。就杨属而言,Howard等[7]对美国窄叶杨(Populus angustifolia)和大叶钻天杨(P. balsamifera)叶片气孔导度的观测结果显示:灌水条件下前者夜间gs在45~308 mmol/(m2·s)之间,后者在37~188 mmol/(m2·s)之间。胡杨夜间气孔导度和蒸腾速率略低于前者而与后者较为接近,远大于杨属植物的最小叶片导度(5 mmol/(m2·s)[7]。胡杨夜间气孔导度和蒸腾速率占白天比例分别为26%和17%,这与美国西部荒漠河岸林(10%~28%)[3]、北美荒漠灌木树种(6%~26%)[5]及美国河岸窄叶杨和大叶钻天杨气孔导度(7.5%~31.3%)[7]大致相当。总之,叶片显微结构观察到夜间气孔的不完全关闭(图 5)为胡杨夜间蒸腾提供了可能,夜间高的气孔导度和蒸腾速率(图 4)为胡杨夜间蒸腾存在提供了直接证据。
关于夜间液流究竟是蒸腾还是组织补水一直存在争议。一些学者认为:相比于白天,夜间液流很弱,且与水汽压差等气象因子无显著相关性,因此夜间液流是组织补水[2, 24-27]。另一些学者认为:夜间液流速率与水汽压差呈显著的正相关,因此把夜间液流归因于夜间蒸腾而非组织补水[4, 23, 28-32]。在我国,学者对干旱区梭梭(Haloxylon ammodendron)、白刺(Nitraria Tangutorum)和沙拐枣(Calligonum mongolicum)3种灌木[33]、华北落叶松(Larix principis-rupprechtii)[34]、广州地区荷木(Schima superba)[35]、华南马占相思树(Acacia mangium)[25]的研究表明:夜间液流与水汽压差、风速等气象因子不相关,因此认为夜间液流是组织补水;而对半干旱区旱柳(Salix matsudana)和小叶杨(Populus simonii)研究表明:夜间液流与水汽压差和风速呈显著相关,认为夜间液流是由组织补水和蒸腾共同造成的[36]。由此可见,夜间液流究竟是蒸腾还是组织补水取决于其是否受水汽压差等气象因子影响,甚至有研究基于此将夜间液流分割为夜间蒸腾和组织补水[13]。
按照SPAC系统理论,水汽压差影响着植物气孔的闭合,从而控制着植物的蒸腾过程。白天,木质部液流速率表征树木蒸腾量,主要受水汽压差影响是无可厚非的[37-38],这与白天液流速率与水汽压差显著相关结果一致(图 3)。但是,夜间平均液流速率为(16.5±5.0)cm/h,而水汽压差可以解释其变化的55%(图 3b),这表明夜间液流由蒸腾和组织补水两部分组成,且应以夜间蒸腾为主。但值得注意的是,胡杨夜间液流速率与风速无关,该结果与加利福尼亚灌木和乔木[39]及澳大利亚温带常绿乔木林[14]一致,而与人工桉树(Populus simonii)林[13]相反。同时,与已有研究相比[4, 30, 39],胡杨夜间液流速率与水汽压差关系并非是线性的,这可能与高的夜间水汽压差限制有关,即高的夜间水汽压差同白天一样限制气孔的开放[2, 14]。值得注意的是,夜间水汽压差仅能解释55%的液流速率变化而非全部,这表明胡杨夜间液流可能还受到其他树木生理过程调节,比如昼夜节律[40]。另外,未来气候变化条件下,随着温度的升高和降水的减少,尤其是夜间(图 1),水汽压差会显著增加,这可能进一步导致夜间蒸腾增加。因此,准确量化夜间蒸腾大小,深刻理解夜间蒸腾发生条件、过程及机理具有重要意义。
4.2 夜间蒸腾发生的时间
关于夜间液流究竟是夜间蒸腾还是组织补水的争论实质是夜间蒸腾何时发生,目前普遍的观点是其发生在午夜之后。例如,对季节性干旱的热带山地多雾林采用组织再填充预测法估算的夜间蒸腾发生时间为01:00[31],而对温带常绿林不同水分利用策略的树种采用基部和树冠液流同步对比确定的夜间蒸腾发生时间为23:00[14],对澳大利亚半干旱区巨桉(Eucalyptus grandis)的观测表明夜间蒸腾发生时间为22:00[30]。上述研究的共同点是夜间VPD普遍较低(0~2 kPa),甚至在前半夜3~4 h内为0 kPa[31],而后半夜液流随水汽压差增加而增加,即这部分增加的液流为夜间蒸腾。
与上述观点不同,笔者认为:夜间高水汽压差条件下,胡杨夜间液流一直存在,但黎明前树干液流的突然增加是组织补水。原因是:第1,假设夜间液流的下降是组织补水,采用趋势预测法(图 2,7月27日)计算的液流速率仍高于“真实零值”,同时从液流下降到第1次液流的上升是与水汽压差变化一致的;而第2次上升(02:00以后)是与水汽压差下降相反的。第2,叶片气孔从完全开放到部分关闭乃至完全关闭是一个渐变的过程,其叶片蒸腾速率也是逐渐递减的(图 4,5),因此夜间气孔不会突然开放以释放水分。第3,基于对液流速率与径向生长(Rd)和木质部水势(Ψs)的同步观测(图 6)表明:在夜间第2次Vs增加的同时,Rd和Ψs也是增加的,即增加的液流导致木质部含水量增加、木质部扩张,说明这一时段是组织补水。
![]() 图 6 胡杨径向生长(Rd)、木质部水势(Ψs)与液流速率(Vs)的变化(2015年6月17—18日)a.径向生长Radial growth; b.木质部水势Stem water potential; c.液流速率Sap velocityFigure 6. Changes of radial growth (Rd), stem water potential (Ψs) and sap velocity (Vs) of P. euphratica during June 17 to 18, 2015
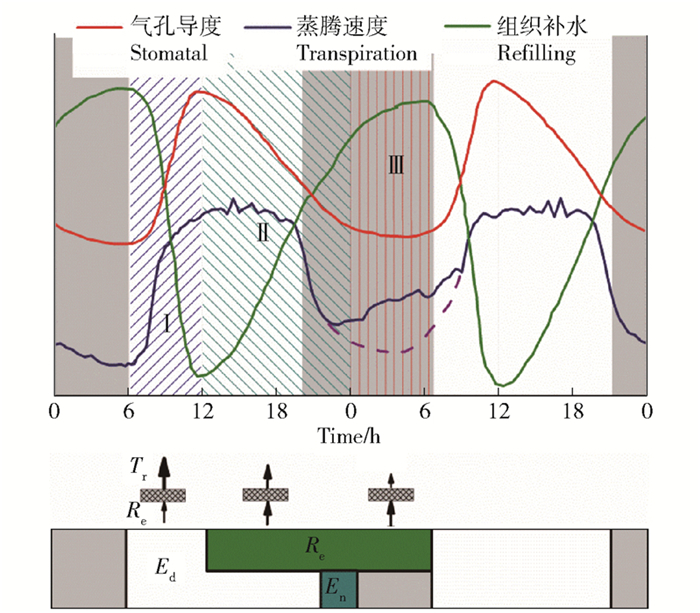
图 6 胡杨径向生长(Rd)、木质部水势(Ψs)与液流速率(Vs)的变化(2015年6月17—18日)a.径向生长Radial growth; b.木质部水势Stem water potential; c.液流速率Sap velocityFigure 6. Changes of radial growth (Rd), stem water potential (Ψs) and sap velocity (Vs) of P. euphratica during June 17 to 18, 2015综上,植物体的蒸腾失水和组织补水是处于动态平衡的,蒸腾失水和组织补水的关系大致可以划分为3个阶段(图 7)。第Ⅰ阶段:从早晨至正午前后,随着蒸腾拉力(水汽压差)的增加,叶片气孔导度和蒸腾速率增大(图 4),树干液流速率增加(图 2),木质部水势下降(图 6),组织补水远远小于蒸腾失水,这一阶段受蒸腾失水控制植物体内的水分是减少的。第Ⅱ阶段,从正午至午夜,虽然蒸腾拉力仍在增加,但气孔导度和蒸腾速率开始下降,木质部水势增加,组织补水开始直至第2天黎明,并与蒸腾维持相对平衡的状态,表现为树干液流稳定变化,存在夜间蒸腾。第Ⅲ阶段,从午夜至黎明,蒸腾拉力很小,气孔部分关闭,木质部水势和夜间液流均增加,组织补水远远大于部分气孔开放引起的失水,说明增加的水分主要是对白天蒸腾失水的补充,这一阶段是完全的组织补水。由此可见,夜间液流变化受夜间蒸腾和组织补水共同影响,并不是相互排斥的。
![]() 图 7 胡杨木质部蒸腾失水与组织补水关系红线为气孔导度;蓝线为蒸腾速率(Tr),包括白天蒸腾(Ed)和夜间蒸腾(En);绿线为组织补水(Refilling, Re)。下图箭头大小代表蒸腾失水与组织补水的相对值。Figure 7. Relationship between transpiration and tissue refilling for the xylem of P. euphraticaIn which red line is stomatal conductance, blue line is transpiration (Tr), including daytime transpiration (Ed) and night-time transpiration (En), and green line is tissue refilling (Re). The size of arrow below represent the relative magnitude of Tr and Re.
图 7 胡杨木质部蒸腾失水与组织补水关系红线为气孔导度;蓝线为蒸腾速率(Tr),包括白天蒸腾(Ed)和夜间蒸腾(En);绿线为组织补水(Refilling, Re)。下图箭头大小代表蒸腾失水与组织补水的相对值。Figure 7. Relationship between transpiration and tissue refilling for the xylem of P. euphraticaIn which red line is stomatal conductance, blue line is transpiration (Tr), including daytime transpiration (Ed) and night-time transpiration (En), and green line is tissue refilling (Re). The size of arrow below represent the relative magnitude of Tr and Re.5. 结论
综上,本文采用木质部液流、叶片气体交换及显微结构观测首次证实胡杨存在夜间蒸腾。胡杨夜间叶片气孔处于不完全关闭状态,气孔导度和蒸腾速率均不为0,且分别占白天的26%和17%;平均液流速率白天为31.3cm/h,夜间为16.5cm/h,夜间约为白天的53%,无论是白天还是夜间,液流速率与水汽压差均呈显著的对数关系,水汽压差可以解释55%的夜间液流变化,说明夜间液流由蒸腾和组织补水两部分组成。值得注意的是,午夜后夜间液流的突然增加是组织补水而非夜间蒸腾,即夜间蒸腾主要发生在日落后至午夜,这表明夜间蒸腾能够促进组织补水以有效缓解植物水分亏缺。未来,如何将夜间液流分割为夜间蒸腾和组织补水,并提高对夜间蒸腾发生条件、过程及机理的理解还有待深入。
-
![]()
图 1 额济纳气象站1961—2012年日平均(Mean)、最高(Max.)、最低(Min.)气温(Ta)及相对湿度(RH)变化
Figure 1. Changes of daily mean, maximum, minimum air temperature (Ta) and relative humidity (RH) in 1961-2012 from Ejin Meteorological Station
![]()
图 2 胡杨树干液流速率(Vs)及水汽压差(VPD)日变化
2012年7月25—28日,图中阴影部分表示夜间。
Figure 2. Daily variation of sap flow velocity (Vs) of Populus euphratica and VPD
The shaded area represent the night-time (25-28, July 2012).
![]()
图 3 胡杨2012年生长季白天和夜间平均液流速率(Vs)变化及其与水汽压差(VPD)的关系
Figure 3. Variations of mean Vs of daytime and night-time and the relationship between Vs and VPD during the growing season in 2012 for P. euphratica
![]()
图 4 胡杨叶片气孔导度(gs)和蒸腾速率(Tr)典型日变化(2014年7月17—18日,7月31日—8月1日和9月2—3日)
Figure 4. Typical daily variations of stomatal conductance (gs) and transpiration rate (Tr) of P. euphratica during July 17 to 18, July 31 to August 1 and September 2 to 3, 2014
![]()
图 5 胡杨幼树白天和夜间叶片气孔显微结构变化(2015年7月1—2日)
Figure 5. Changes of leaf stomatal microstructure of P. euphratica at daytime and nighttime during July 1 to 2, 2015
![]()
图 6 胡杨径向生长(Rd)、木质部水势(Ψs)与液流速率(Vs)的变化(2015年6月17—18日)
a.径向生长Radial growth; b.木质部水势Stem water potential; c.液流速率Sap velocity
Figure 6. Changes of radial growth (Rd), stem water potential (Ψs) and sap velocity (Vs) of P. euphratica during June 17 to 18, 2015
![]()
图 7 胡杨木质部蒸腾失水与组织补水关系
红线为气孔导度;蓝线为蒸腾速率(Tr),包括白天蒸腾(Ed)和夜间蒸腾(En);绿线为组织补水(Refilling, Re)。下图箭头大小代表蒸腾失水与组织补水的相对值。
Figure 7. Relationship between transpiration and tissue refilling for the xylem of P. euphratica
In which red line is stomatal conductance, blue line is transpiration (Tr), including daytime transpiration (Ed) and night-time transpiration (En), and green line is tissue refilling (Re). The size of arrow below represent the relative magnitude of Tr and Re.
表 1 试验期内额济纳站气象要素变化
Table 1 Changes in meteorological factors in Ejin Meteorological Station during experiment period
年份
Year降水量
Precipitation (P)/mm日照时间
Sunshine duration(N)/h气温
Air temperature(Ta)/℃相对湿度
Relative humidity(RH)/%水汽压差
Vapor pressure difference (VPD)/kPa风速
Wind speed(U)/(m·s-1)2012 32.7 8.77±3.04 9.44±14.47 27.72±12.23 1.66±1.27 2.60±1.09 2014 17.2 8.33±3.15 10.55±13.30 30.16±12.14 1.70±1.21 2.63±1.11 2015 44.4 8.63±3.07 10.66±12.78 32.05±14.58 1.71±1.22 2.68±1.11 Sig. No No Yes No No  下载: 导出CSV
下载: 导出CSV
-
[1] RAWSON H M, CLARKE J M. Nocturnal transpiration in wheat[J]. Australian Journal of Plant Physiology, 1988, 15: 397-406.
[2] CAIRD M A, RICHARDS J H, DONOVAN L A. Nighttime stomatal conductance and transpiration in C3 and C4 plants[J]. Plant Physiology, 2007, 143(1): 4-10. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=PubMed000001681649
[3] SNYDER K A, RICHARDS J H, DONOVAN L A. Night-time conductance in C3 and C4 species: do plants lose water at night?[J]. Journal of Experiment Botany, 2003, 54: 861-865. doi: 10.1093/jxb/erg082
[4] DAWSON T E, BURGESS S S O, TU K P, et al. Nighttime transpiration in woody plants from contrasting ecosystems[J]. Tree Physiology, 2007, 27: 561-575. doi: 10.1093/treephys/27.4.561
[5] OGLE K, LUCAS R W, BENTLEY L P, et al. Differential daytime and night-time stomatal behavior in plants from North American deserts[J]. New Phytologist, 2012, 194(2): 464-476. doi: 10.1111/j.1469-8137.2012.04068.x
[6] MOORE G W, CLEVERLY J R, OWENS M K. Nocturnal transpiration in riparian Tamarix thickets authenticated by sap flux, eddy covariance and leaf gas exchange measurements[J]. Tree Physiology, 2008, 28: 521-528. doi: 10.1093/treephys/28.4.521
[7] HOWARD A R, DONOVAN L A. Soil nitrogen limitation does not impact nighttime water loss in Populus[J]. Tree Physiology, 2010, 30(1): 23-31. https://www.ncbi.nlm.nih.gov/pubmed/19959599
[8] DAMIÁN C, MARÍA A E, VICTOR J L, et al. Populus species from diverse habitats maintain high night-time conductance under drought[J]. Tree Physiology, 2016, 36(2): 229-242. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=84d9f60cc24ba0df2777b42dce988b40
[9] YU T F, FENG Q, SI J H, et al. Tamarix ramosissima stand evapotranspiration and its association with hydroclimatic factors in an arid region in northwest China[J]. Journal of Arid Environments, 2017, 138: 18-26. doi: 10.1016/j.jaridenv.2016.11.006
[10] 司建华, 冯起, 鱼腾飞, 等.植物夜间蒸腾及其生态水文效应研究进展[J].水科学进展, 2014, 25(6): 907-914. http://d.old.wanfangdata.com.cn/Periodical/skxjz201406017 SI J H, FENG Q, YU T F, et al. Research advances in nighttime transpiration and its eco-hydrological implications[J]. Advances in Water Science, 2014, 25(6): 907-914. http://d.old.wanfangdata.com.cn/Periodical/skxjz201406017
[11] NADEZHDINA N. Sap flow index as an indicator of plant water status[J]. Tree Physiology, 1999, 19: 885-891. doi: 10.1093/treephys/19.13.885
[12] BURGESS S S O, ADAMS M A, TURNER N C, et al. An improved heat pulse method to measure low and reverse rates of sap flow in woody plants[J]. Tree Physiology, 2001, 21: 589-598. doi: 10.1093/treephys/21.9.589
[13] ZEPPEL M J, LEWIS J D, MEDLYN B, et al. Interactive effects of elevated CO2 and drought on nocturnal water fluxes in Eucalyptus saligna[J]. Tree Physiology, 2011, 31(9): 932-944. doi: 10.1093/treephys/tpr024
[14] PHILLIPS N G, LEWIS J D, LOGAN B A, et al. Inter- and intra-specific variation in nocturnal water transport in Eucalyptus[J]. Tree Physiology, 2010, 30: 586-596. doi: 10.1093/treephys/tpq009
[15] YU T F, FENG Q, SI J H, et al. Hydraulic redistribution of soil water by roots of two desert riparian phreatophytes in northwest China's extremely arid region[J]. Plant and Soil, 2013, 372: 297-308. doi: 10.1007/s11104-013-1727-8
[16] IPCC. Climate change 2007: the physical science basis[M]. Cambridge: Cambridge University Press, 2007.
[17] 鱼腾飞, 冯起, 司建华, 等.胡杨根系水力再分配的模式、大小及其影响因子[J].北京林业大学学报, 2014, 36(2): 22-29. http://j.bjfu.edu.cn/article/id/9977 YU T F, FENG Q, SI J H, et al. Patterns, magnitude and controlling factors of hydraulic redistribution by Populus euphratica roots[J]. Journal of Beijing Forestry University, 2014, 36(2) 22-29. http://j.bjfu.edu.cn/article/id/9977
[18] ALLEN R G, PEREIRA L S, RAES D. Crop evapotranspiration: guidelines for computing crop water requirements: FAO irrigation and drainage paper 56[C]. Rome: FAO, 1998.
[19] YANG Y T, GUAN H D, HUTSON J L, et al. Examination and parameterization of the root water uptake model from stem water potential and sap flow measurements[J]. Hydrological Processes, 2013, 27: 2857-2863. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=10.1002/hyp.9406
[20] 赵传燕, 赵阳, 彭守璋, 等.黑河下游绿洲胡杨生长状况与叶生态特征[J].生态学报, 2014, 34(16): 4518-4525. http://d.old.wanfangdata.com.cn/Periodical/stxb201416007 ZHAO C Y, ZHAO Y, PENG S Z, et al. The growth state of Populus euphratica Oliv. and its leaf ecological characteristics in the lower reaches of Heihe River[J]. Acta Ecologica Sinica, 2014, 34(16): 4518-4525. http://d.old.wanfangdata.com.cn/Periodical/stxb201416007
[21] BURGESS S S O, DAWSON T E. The contribution of fog to the water relations of Sequoia sempervirens (D. Don): foliar uptake and prevention of dehydration[J]. Plant, Cell and Environment, 2004, 27: 1023-1034. doi: 10.1111/j.1365-3040.2004.01207.x
[22] GOLDSMITH G R. Changing directions: the atmosphere-plant-soil continuum[J]. New Phytologist, 2013, 199(1): 4-6. doi: 10.1111/nph.12332
[23] DALEY M J, PHILLIPS N G. Interspecific variation in nighttime transpiration and stomatal conductance in a mixed New England deciduous forest[J]. Tree Physiology, 2006, 26: 411-419. doi: 10.1093/treephys/26.4.411
[24] GOLDSTEIN G, ANDRADE J L, MEINZER F C, et al. Stem water storage and diurnal patterns of water use in tropical forest[J]. Plant, Cell and Environment, 1998, 21: 397-406. doi: 10.1046/j.1365-3040.1998.00273.x
[25] WANG H, ZHAO P, WANG Q, et al. Nocturnal sap flow characteristics and stem water recharge of Acacia mangium[J]. Frontiers of Forestry in China, 2008, 3(1): 72-78. doi: 10.1007/s11461-008-0005-z
[26] OREN R, PHILLIPS N G, EWERS B E, et al. Sap-flux-scaled transpiration responses to light, vapor pressure deficit, and leaf area reduction in a flooded Taxodium distichum forest[J]. Tree Physiology, 1999, 19: 337-347. doi: 10.1093/treephys/19.6.337
[27] PHILLIPS N G, RYAN M G, BOND B J, et al. Reliance on stored water increases with tree size in three species in the Pacific Northwest[J]. Tree Physiology, 2003, 23: 237-245. doi: 10.1093/treephys/23.4.237
[28] KAVANAGH K L, PANGLE R P, SCHOTZKO A D. Nocturnal transpiration causing disequilibrium between soil and stem predawn water potential in mixed conifer forests of Idaho[J]. Tree Physiology, 2007, 27: 621-629. doi: 10.1093/treephys/27.4.621
[29] HOGG E H, HURDLE P A. Sap flow in trembling aspen implications for stomatal responses to vapor pressure deficit[J]. Tree Physiology, 1997, 17: 501-509. doi: 10.1093/treephys/17.8-9.501
[30] BENYON R G. Nighttime water use in an irrigated Eucalyptus grandis plantation[J]. Tree Physiology, 1999, 19: 853-859. doi: 10.1093/treephys/19.13.853
[31] ALVARADO-BARRIENTOS M S, HOLWERDA F, GEISSERT D R, et al. Nighttime transpiration in a seasonally dry tropical montane cloud forest environment[J]. Trees, 2014, 29(1): 259-274. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=f9c209cebdb64fb217cb137aafdff8d3
[32] BUCKLEY T N, TURNBULL T L, PFAUTSCH S, et al. Nocturnal water loss in mature subalpine Eucalyptus delegatensis tall open forests and adjacent E. pauciflora woodlands[J]. Ecology Evolution, 2011, 1(3): 435-450. doi: 10.1002/ece3.44
[33] 徐世琴, 吉喜斌, 金博文.西北干旱区典型固沙植物夜间耗水及其影响因素[J].西北植物学报, 2015, 35(7): 1443-1450. http://d.old.wanfangdata.com.cn/Periodical/xbzwxb201507022 XU S Q, JI X B, JIN B W. Nighttime water use and its influencing factors for typical sand binding plants in the arid region of northwest China[J]. Acta Botanica Boreali-Occidentalia Sinica, 2015, 35(7): 1443-1450. http://d.old.wanfangdata.com.cn/Periodical/xbzwxb201507022
[34] 王艳兵, 德永军, 熊伟, 等.华北落叶松夜间树干液流特征及生长季补水格局[J].生态学报, 2013, 33(5): 1375-1385. http://d.old.wanfangdata.com.cn/Periodical/stxb201305005 WANG Y B, DE Y J, XIONG W, et al. The characteristics of nocturnal sap flow and stem water recharge pattern in growing season for a Larix principis-rupprechtii plantation[J]. Acta Ecologica Sinica, 2013, 33(5): 1375-1385. http://d.old.wanfangdata.com.cn/Periodical/stxb201305005
[35] 周翠鸣, 赵平, 倪广艳, 等.广州地区荷木夜间树干液流补水的影响因子及其对蒸腾的贡献[J].应用生态学报, 2012, 23(7): 1751-1757. http://d.old.wanfangdata.com.cn/Periodical/yystxb201207003 ZHOU C M, ZHAO P, NI G Y, et al. Water recharge through nighttime stem sap flow of Schima superba in Guangzhou Region of Guangdong Province, South China: affecting factors and contribution to transpiration[J]. Chinese Journal of Applied Ecology, 2012, 23(7): 1751-1757. http://d.old.wanfangdata.com.cn/Periodical/yystxb201207003
[36] 尹立河, 黄金廷, 王晓勇, 等.陕西榆林地区旱柳和小叶杨夜间树干液流变化特征分析[J].西北农林科技大学学报(自然科学版), 2013, 41(8): 85-90. http://d.old.wanfangdata.com.cn/Periodical/xbnydxxb201308014 YIN L H, HUANG J T, WANG X Y, et al. Characteristice of nighttime sap flow of Salix matsudana and Populus simonii in Yulin, Shaanxi[J]. Journal of Northwest A & F University (Natural Science Edition), 2013, 41(8): 85-90. http://d.old.wanfangdata.com.cn/Periodical/xbnydxxb201308014
[37] 赵春彦, 司建华, 冯起, 等.胡杨(Populus euphratica)树干液流特征及其与环境因子的关系[J].中国沙漠, 2014, 34(3): 718-724. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=zgsm201403014 ZHAO C Y, SI J H, FENG Q, et al. Xylem sap flow of Populus euphratica in relation to environmental factors in the lower reaches of Heihe River[J]. Journal of Desert Research, 2014, 34(3): 718-724. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=zgsm201403014
[38] 司建华, 冯起, 张小由, 等.极端干旱区荒漠河岸林胡杨生长季树干液流变化[J].中国沙漠, 2007, 27(3): 442-447. doi: 10.3321/j.issn:1000-694X.2007.03.016 SI J H, FENG Q, ZHANG X Y, et al. Sap flow of Populus euphratica in desert riparian forest in extreme arid region during the growing season[J]. Journal of Desert Research, 2007, 27(3): 442-447. doi: 10.3321/j.issn:1000-694X.2007.03.016
[39] FISHER J B, BALDOCCHI D D, MISSON L, et al. What the towers don't see at night: nocturnal sap flow in trees and shrubs at two AmeriFlux sites in California[J]. Tree Physiology, 2007, 27: 597-610. doi: 10.1093/treephys/27.4.597
[40] RESCO DE DIOS V, DIAZ-SIERRA R, GOULDEN M L, et al. Woody clockworks: circadian regulation of night-time water use in Eucalyptus globulus[J]. New Phytologist, 2013, 200(3): 743-752. doi: 10.1111/nph.12382
-
期刊类型引用(3)
1. 李潇,王汉时,王宏星,蒋路平,庞忠义,彭彦辉,赵曦阳. 灌溉和施肥对‘新林1号’杨生长和光合生理特性的影响. 植物研究. 2025(01): 77-87 .  百度学术
百度学术
2. 张聪,贾炜玮,郭昊天,范迎新. 不同施肥措施的人工落叶松生长差异性. 东北林业大学学报. 2024(11): 1-9 . 百度学术
3. 王亚飞,贺曰林,杨红青,祝维,贾黎明,席本野. 灌溉施肥对杨树人工林林木及地力效应研究进展. 世界林业研究. 2023(05): 63-69 . 百度学术
其他类型引用(0)




计量
- 文章访问数: 2199
- HTML全文浏览量: 522
- PDF下载量: 55
- 被引次数: 3
