
Study on interspecific associations of typical mid-subtropical natural broadleaved forest based on stratification
-
摘要: 亚热带天然常绿阔叶林群落的乔木层普遍具有自然成层特征,通过划分林层,研究了两个典型中亚热带天然阔叶林群落各层组成种间的种间联结,以揭示各层林木间的关系。结果表明:两个群落树种丰富,乔木层分别有65和53个树种;乔木层都可划分为受光层和非受光层两个亚层,受光层对群落起控制作用;两个群落的受光层所有树种都是总体负关联,导致了乔木层的总体负关联;无论是乔木层还是两个亚层,各层林木均可结成数量可观的成对组合,但大多数树种两两之间关系松散或相互独立,只有个别树种与其他树种有显著的正或负关联,表明调查群落主要受随机过程控制。在生物多样性保护实践中需要关注猴欢喜和新木姜子,因为在包含这两个种的显著关联的种对中,前者与其他树种的均为负关联,后者则以正关联为主。Abstract: Stratification of arbor layer is popular in subtropical natural evergreen broadleaved forests. Two typical natural broadleaved forest communities in mid-subtropical zone were studied to reveal the interspecific associations of arbor layer and its sub-layers. There were 65 and 53 species in the arbor layer of the two communities, respectively.Arbor layer of the two communities could be divided into two sub-layers, light receiving layer (LRL) and non-light receiving layer (NLRL), and LRL dominated the communities. All species in LRL of two communities were totally negative association, which resulted in the totally negative associationin of arbor layer.Whether arbor layer or the two sub-layers, species in each layer could form numerous combinations in pair-wise approach, but most species tended to be independent with each other and few species were significantly positive or negative associated with other species, suggesting that random processes dominated the two communities. Attention should be paid to Sloanea sinensis and Neolitsea aurata in biodiversity protection practice since the former was always negatively associated with other species, while the later was mainly positively associated with other species among those species pairs with significant association.
-
紫薇(Lagerstroemia indica)为千屈菜科(Lythraceae)紫薇属落叶灌木或小乔木,具有树姿优美、花色艳丽、花期长等特点,是我国乃至世界有名的夏季观花小乔木,具有很高的观赏价值。育种工作者培育出的紫叶紫薇品种(Lagerstroemia indica ‘Ebony Ember’),其叶色始终保持紫红色,进一步增加了紫薇的观赏价值。
植物叶片的呈色机理复杂。在许多彩叶植物上的研究表明,叶片色素的种类、比值及分布决定了叶片的呈色[1]。叶片的呈色色素主要为叶绿素、类胡萝卜素和花青苷,其中花青苷主要呈现红色到蓝色一系列颜色,在红(紫)叶植物叶片呈色中起主导作用[2-4]。朱书香等[5]发现李属(Prunus)植物叶片随着花青苷含量比例的高—低—高变化呈现红—绿—红的过渡。蔡雪雁等[6]发现紫叶桃(Prunus persica f. atropurpurea)叶片中花青苷是叶片呈现红色的主要色素,而叶绿素和类胡萝卜素对叶片红色的加深起到了重要作用。马艳芝[7]以4种红色叶植物的叶片为材料,观察在叶片不同结构中色素的分布情况,发现由于色素分布部位不同导致叶片呈现出不同程度的红色或紫红色,并且叶片上下表面颜色也有所不同。可溶性糖不仅参与花青苷的合成,还作为一种信号调控花青苷合成[8]。研究表明,可溶性糖可以影响多种植物花青苷的积累[9-10]。
叶片的不同呈色影响叶片的光合特性,与叶片中色素的变化有关[11]。前人对于红叶植物光合特性的研究结论并不一致,‘Rutger’桃(Prunus persica f. atropurpurea ‘Rutger’)(红叶)的净光合速率、表观量子效率、羧化效率显著低于白芒蟠桃(Prunus persica f. compressa ‘Baimangpantao’)(绿叶)[12]。李属4种红叶树种净光合速率、实际光化学效率、光化学淬灭系数均低于绿叶树种,而非光化学淬灭系数高于绿叶树种,热耗散能力较强[13]。美国红栌(Cotinus coggygria ‘Purpureus’)的光合速率和水分利用效率在各个季节都大于普通黄栌(Cotinus coggygria),并且年光合能力也大于普通黄栌[14]。另外,一品红(Euphorbia pulcherrima)的红叶与绿叶中色素组成差异巨大,但在光合能力上并未有显著差异[15]。
目前,国内外对于紫薇的研究主要集中在种质资源、育种、抗病性、花色、株型、药用价值等方面[16-19],对叶色的研究甚少。本研究通过比较紫薇绿叶品种‘Arapaho’和紫叶品种‘Ebony Embers’叶片的色素含量、可溶性糖含量、色素分布及净光合速率等生理生化指标,分析紫叶紫薇的呈色机理和光合特性,旨在为选育紫薇优良彩叶品种及优化紫薇的园林栽培配置提供理论基础。
1. 材料与方法
1.1 试验材料
试验样品于2016年5—9月取自国家花卉工程技术研究中心小汤山基地。材料为4年生紫叶紫薇品种和绿叶紫薇品种(L. indica ‘Arapaho’),选取生长健壮、长势一致的植株。从2016年5月6日起,于叶片萌发后第5天(S1)、第10天(S2)、第15天(S3)、第20天(S4)、第30天(S5)分别选择向阳部位生长健康的叶片取样,保存于-80 ℃。
1.2 试验方法
1.2.1 光合色素含量测定
按照朱广廉等[20]的方法并改进,取0.1 g叶片研磨,用80%丙酮溶液进行叶绿素和类胡萝卜素的提取,用紫外分光光度计(Biomate 3S)在470、645、662 nm下测定OD值,按以下公式计算各光合色素的含量:
Ca=(12.7OD663−2.69OD645)V/M Cb=(22.9OD645−4.68OD663)V/M Ca+b=Ca+Cb Cx=1000OD470−3.27Ca−Cb)V/229M 式中:Ca代表叶绿素a的含量(mg/g),Cb代表叶绿素b的含量(mg/g),Ca+b代表叶绿素总含量(mg/g),Cx代表类胡萝卜素的含量(mg/g),V代表提取液体积(L),M代表叶片鲜质量(g)。
1.2.2 总花青苷含量测定
按照何奕昆等[21]的方法并改进,取0.1 g叶片研磨,用体积分数0.1%的盐酸甲醇溶液浸提花青苷,使用紫外分光光度计(Biomate 3S)在532、650 nm下测定OD值,按以下公式计算花青苷的含量:
Cant=(OD532−0.25OD650)V/0.0462M 式中:Cant代表花青苷含量(mg/g)。
1.2.3 可溶性糖含量测定
采用蒽酮比色法于630 nm波长处测定OD值,取标准糖溶液稀释成0~100 μg/mL不同质量浓度的溶液,测定OD值后绘制标准曲线,根据标准曲线计算出可溶性糖含量[22]。
1.2.4 切片制备
参照李亚蒙等[23]的方法,选取新长成的功能叶,进行徒手切片,采用双刀片紧压并排碎切法,切取叶片中部带叶脉的部分制成切片进行观察拍照。
1.2.5 光合作用日变化测定
从2个品种紫薇中各选取3株,每株测定向阳面位置相同的3片成熟叶片,每片叶重复3次,于2015年9月1—3日,选择晴朗无云的天气,从07:00开始,每隔2 h测定一次,直到17:00。测定时利用自然光照和大气CO2,用LI-6400便携式光合测定系统测定净光合速率(Pn)、气孔导度(Gs)、胞间二氧化碳浓度(Ci)、蒸腾速率(Tr)。
1.2.6 光响应曲线和CO2响应曲线测定
光响应曲线的测定:从2个品种紫薇中各选取3株,每株测定向阳面位置相同的3片成熟叶片,于2015年9月1—3日,选择天气晴朗的08:00—11:00,用LI-6400便携式光合测定系统控制叶片温度为25 ℃,叶室CO2浓度为400 μmol/mol,设置叶室光照强度从高到低为2 000、1 800、1 500、1 200、1 000、800、500、200、180、150、120、100、80、50、0 μmol/(m2·s),测定Pn,绘制光响应曲线。
CO2浓度响应曲线的测定:从2个品种紫薇中各选取3株,每株测定向阳面位置相同的3片成熟叶片,于2015年9月1—3日,选择天气晴朗的08:00—11:00,用LI-6400便携式光合测定系统控制光照强度为1 700 μmol/(m2·s)(光饱和点附近),设置叶室CO2浓度从低到高为50、100、150、200、300、400、600、800、1 000、1 200、1 500、1 800、2 000 μmol/mol,测定Pn,绘制CO2浓度响应曲线。
1.2.7 叶绿素荧光参数测定
从2个品种紫薇中各选取3株,每株测定向阳面位置相同的3片成熟叶片,用PAM-2500便携式叶绿素荧光仪测定。测定前叶片进行20 min暗适应,测定初始荧光(F0)、最大荧光(Fm)、可变荧光(Fv)、PSⅡ最大光化学效率(Fv/Fm)、PSⅡ潜在光化学活性(Fv/F0)、PSⅡ电子传递量子效率(ΦPSⅡ)、表观光合电子传递速率(ETR)、光化学猝灭系数(qP)、非光化学猝灭系数(qN和NPQ)。
1.3 数据分析
利用SPSS 19.0软件对数据进行差异显著性分析和相关性分析;利用Microsoft Excel 2007作图。
2. 结果与分析
2.1 不同发育时期叶片色素含量变化
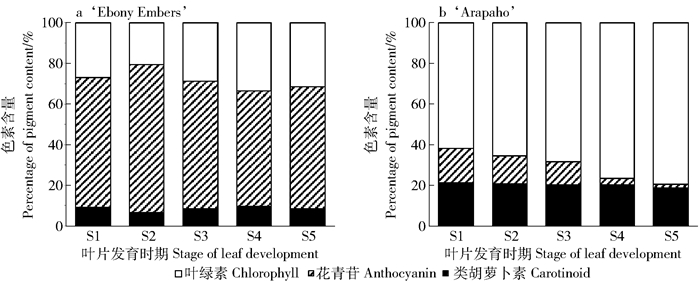
由图 1可知,随着叶片的发育,‘Ebony Embers’始终保持紫色,而‘Arapaho’叶片嫩叶泛红,然后逐渐变为深绿色。‘Ebony Embers’不同发育期叶片的叶绿素和类胡萝卜素含量呈先降低后升高的趋势,除S2时期,其他时期均显著高于绿叶品种,其成熟叶片叶绿素和类胡萝卜素含量分别为2.88和0.79 mg/g,分别是‘Arapaho’的1.35倍和1.55倍;‘Ebony Embers’叶片花青苷含量随着叶片发育逐渐升高,而‘Arapaho’叶片的花青苷含量逐渐降低,‘Ebony Embers’的花青苷含量始终显著高于‘Arapaho’,两者成熟叶片的花青苷含量比值最高达到125.1:1(图 2)。
![]() 图 1 紫薇品种‘Ebony Embers’和‘Arapaho’叶片发育过程中叶色变化S1、S2、S3、S4、S5分别代表叶片萌发后第5天、第10天、第15天、第20天、第30天。下同。Figure 1. Leaf color changes of Lagerstroemia indica 'Ebony Embers' and 'Arapaho' during leaf developmentS1, S2, S3, S4, S5 represent 5 days, 10 days, 15 days, 20 days, 30 days after budding, respectively. The same below.
图 1 紫薇品种‘Ebony Embers’和‘Arapaho’叶片发育过程中叶色变化S1、S2、S3、S4、S5分别代表叶片萌发后第5天、第10天、第15天、第20天、第30天。下同。Figure 1. Leaf color changes of Lagerstroemia indica 'Ebony Embers' and 'Arapaho' during leaf developmentS1, S2, S3, S4, S5 represent 5 days, 10 days, 15 days, 20 days, 30 days after budding, respectively. The same below.![]() 图 2 紫薇品种‘Ebony Embers’和‘Arapaho’叶片发育过程中叶片色素含量变化Figure 2. Pigment content in leaves of Lagerstroemia indica 'Ebony Embers' and 'Arapaho' during leaf development
图 2 紫薇品种‘Ebony Embers’和‘Arapaho’叶片发育过程中叶片色素含量变化Figure 2. Pigment content in leaves of Lagerstroemia indica 'Ebony Embers' and 'Arapaho' during leaf development比较紫薇叶片发育过程中3类色素含量(花青苷:类胡萝卜素:叶绿素)百分比的变化(图 3)发现,各类色素百分比存在明显差异。‘Ebony Embers’叶片中花青苷占最大比例且比值变化不大,约为57%~73%;‘Arapaho’叶片中叶绿素所占比例最大且随着叶片成熟逐渐升高,而花青苷所占比值逐渐降低,类胡萝卜素所占比例没有明显变化。
![]() 图 3 紫薇品种‘Ebony Embers’和‘Arapaho’叶片中色素含量Figure 3. Percentage of pigment contents in leaves of Lagerstroemia indica 'Ebony Embers' and 'Arapaho' during leaf development
图 3 紫薇品种‘Ebony Embers’和‘Arapaho’叶片中色素含量Figure 3. Percentage of pigment contents in leaves of Lagerstroemia indica 'Ebony Embers' and 'Arapaho' during leaf development2.2 不同发育时期叶片可溶性糖含量的变化
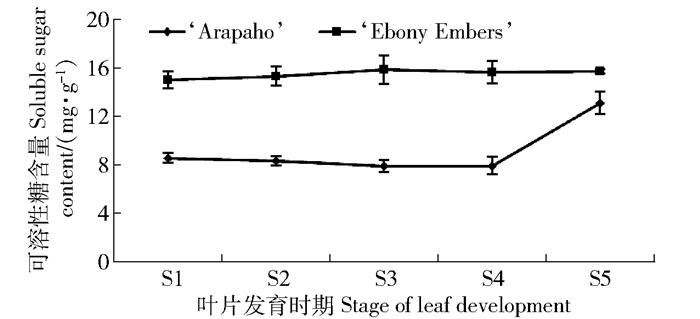
由图 4可知:在叶片发育过程中,‘Arapaho’叶片中可溶性糖含量在前4个时期没有明显的变化,在叶片成熟后,可溶性糖含量迅速升高;而‘Ebony Embers’叶片中可溶性糖含量在叶片发育时期没有明显变化。相关性分析表明,可溶性糖与叶绿素、类胡萝卜素之间没有显著相关性,‘Ebony Embers’中可溶性糖含量与花青苷之间呈显著相关性,相关系数为0.913,而在绿叶品种中则没有显著相关性。
![]() 图 4 紫薇品种‘Ebony Embers’和‘Arapaho’叶片可溶性糖含量变化Figure 4. Changes of soluble sugar content in leaves of Lagerstroemia indica 'Ebony Embers' and 'Arapaho' during leaf development
图 4 紫薇品种‘Ebony Embers’和‘Arapaho’叶片可溶性糖含量变化Figure 4. Changes of soluble sugar content in leaves of Lagerstroemia indica 'Ebony Embers' and 'Arapaho' during leaf development2.3 叶片的色素分布
不同叶色紫薇的叶片结构没有明显差异,只有色素种类与分布的差异(图 5)。‘Ebony Embers’中有明显的花青苷的存在,主要分布在栅栏组织和海绵组织靠近表皮的部分,在显微镜下呈现紫红色;而在‘Arapaho’中,叶绿素分布在栅栏组织和海绵组织,几乎没有花青苷的存在,在显微镜下呈现绿色。
![]() 图 5 紫薇品种‘Ebony Embers’和‘Arapaho’叶片中色素的分布Figure 5. Distribution of pigment in leaves of Lagerstroemia indica 'Ebony Embers' and 'Arapaho' during leaf development
图 5 紫薇品种‘Ebony Embers’和‘Arapaho’叶片中色素的分布Figure 5. Distribution of pigment in leaves of Lagerstroemia indica 'Ebony Embers' and 'Arapaho' during leaf development2.4 光合指标日变化
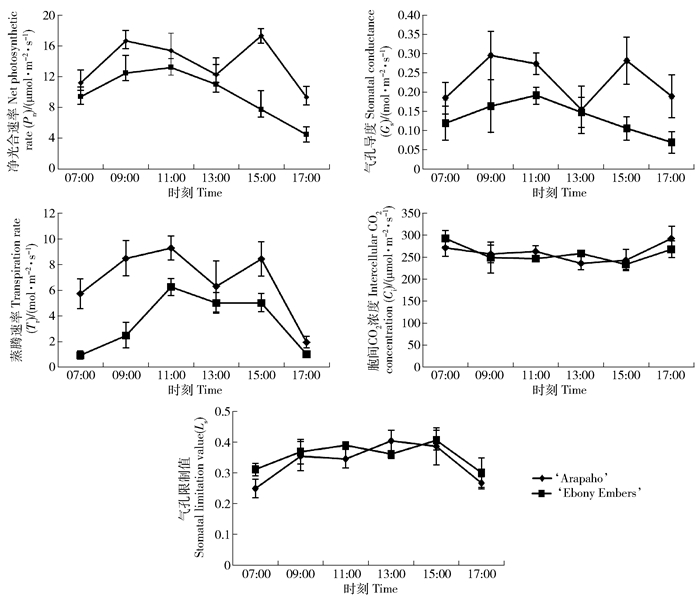
由图 6可知:‘Arapaho’光合速率日变化、气孔导度、蒸腾速率和气孔限制值变化均为双峰曲线,存在明显的光合午休现象,而‘Ebony Embers’除气孔限制值外其他指标变化均为单峰曲线,无光合午休现象;‘Arapaho’胞间CO2浓度呈现“V”变化趋势,而‘Ebony Embers’呈现“W”变化趋势,于13:00出现短暂升高;‘Arapaho’气孔限制值先升高,于13:00达到峰值后降低,而‘Ebony Embers’则于13:00出现短暂下降。一天中‘Arapaho’的净光合速率、气孔导度和蒸腾速率始终高于‘Ebony Embers’,而两者胞间CO2浓度和气孔限制值没有显著差异。
![]() 图 6 紫薇品种‘Ebony Embers’和‘Arapaho’光合参数日变化Figure 6. Diurnal changes of photosynthetic indexes of Lagerstroemia indica 'Ebony Embers' and 'Arapaho'
图 6 紫薇品种‘Ebony Embers’和‘Arapaho’光合参数日变化Figure 6. Diurnal changes of photosynthetic indexes of Lagerstroemia indica 'Ebony Embers' and 'Arapaho'2.5 光合指标相关性分析
净光合速率与气孔导度极显著相关,与蒸腾速率显著相关,气孔导度与蒸腾速率之间相关显著(表 1),再结合图 6可知,‘Arapaho’净光合速率在午休阶段明显下降,Gs和Ci降到最低值,Ls升高到最高值,说明绿叶品种光合午休是气孔因素导致的。
表 1 光合指标间的相关性分析Table 1. Correlation analysis of photosynthetic indexes of Lagerstroemia indica 'Ebony Embers' and 'Arapaho'指标Index Pn Cs Tr Ci Pn 1 Cs 0.954** 1 Tr 0.797** 0.762** 1 Ci -0.276 -0.124 -0.543 1 注:**表示相关极显著(P < 0.01)。Note: ** means highly significant correlation (P < 0.01). 2.6 光合作用光响应和CO2浓度响应曲线
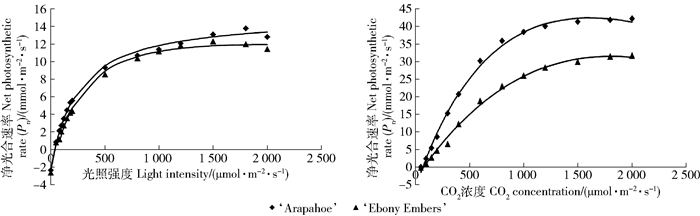
2个品种的光响应曲线和CO2浓度响应曲线相似,但在相同光照强度或CO2浓度下‘Ebony Embers’的净光合速率始终低于‘Arapaho’;‘Arapaho’的光饱和点、CO2饱和点和羧化效率显著高于‘Ebony Embers’(图 7、表 2),说明‘Arapaho’对强光和不同浓度CO2的利用能力较‘Ebony Embers’高。
![]() 图 7 紫薇品种‘Ebony Embers’和‘Arapaho’净光合速率的光、CO2响应曲线Figure 7. Light-response and CO2-response curves of net photosynthetic rate of Lagerstroemia indica 'Ebony Embers' and 'Arapaho'表 2 紫薇品种‘Ebony Embers’和‘Arapaho’的主要光合参数Table 2. Main photosynthetic indexes of Lagerstroemia indica 'Ebony Embers' and 'Arapaho'
图 7 紫薇品种‘Ebony Embers’和‘Arapaho’净光合速率的光、CO2响应曲线Figure 7. Light-response and CO2-response curves of net photosynthetic rate of Lagerstroemia indica 'Ebony Embers' and 'Arapaho'表 2 紫薇品种‘Ebony Embers’和‘Arapaho’的主要光合参数Table 2. Main photosynthetic indexes of Lagerstroemia indica 'Ebony Embers' and 'Arapaho'品种
Cultivar光饱和点Light saturation point/(μmol·m-2·s-1) 光补偿点Light compensation point/(μmol·m-2·s-1) CO2饱和点
CO2 saturation point/(μmol·mol-1)CO2补偿点
CO2 compensation point/(μmol·mol-1)表观量子效率
Apparent quantum/(mol·mol-1)羧化效率
Carboxylation efficiency/(mol·m-2)‘Ebony Ember’ 1 717.67±64.79 37.87±1.56 1 347±50 76.04±5.38 0.033±0.004 6 0.028±0.004 5 ‘Arapaho’ 1 952±75.63* 36.16±0.47 1 550±75* 67.17±4.32 0.038±0.007 6 0.062±0.003** 注:*表示相关显著(P < 0.05),**表示相关极显著(P < 0.01)。下同。Notes: * means significant correlation (P < 0.05), ** means highly significant correlation (P < 0.01). The same below. 2.7 叶绿素荧光参数
由表 3可知:2个品种Fv/Fm和Fv/F0值之间没有显著差异,说明两者之间光系统Ⅱ原初光能转换效率和光系统Ⅱ潜在活性相近;‘Ebony Embers’的ΦPSⅡ值和ETR值显著小于‘Arapaho’,qN和NPQ值显著大于‘Arapaho’,表明‘Arapaho’的PSⅡ电子传递活性高于‘Ebony Embers’,在光能利用中占有优势,而‘Ebony Embers’的热耗散效率高于‘Arapaho’。
表 3 紫薇品种‘Ebony Embers’和‘Arapaho’主要叶绿素荧光参数Table 3. Main chlorophyll fluoresce parameters of Lagerstroemia indica 'Ebony Embers' and 'Arapaho'品种Cultivar Fv/Fm Fv/F0 ΦPSⅡ NPQ qN qP ETR ‘Ebony Ember’ 0.81±0.01 4.13±0.16 0.26±0.03 2.06±0.64* 0.76±0.07* 0.46±0.01 52.6±5.1 ‘Arapaho’ 0.8±0.01 3.99±0.26 0.38±0.06* 1.19±0.25 0.63±0.06 0.59±0.09 75.9±12.9* 3. 讨论
植物色素是植物叶色形成的决定性物质,对于多数红(紫)色叶系植物来说,花青苷含量以及花青苷与叶绿素比值是决定叶色的最主要因素[2-3, 11]。在叶片发育过程中,‘Ebony Embers’光合色素及花青苷含量整体上均高于‘Arapaho’,其中‘Ebony Embers’叶片花青苷含量随着叶片的发育逐渐升高,‘Arapaho’则呈现相反的趋势。另外,2个品种叶片中各色素所占比例的变化与其叶片红色的呈现情况相吻合,故认为花青苷是‘Ebony Embers’叶片呈现紫红色的决定色素,并且花青苷含量比值决定了叶片红色呈现的程度,而叶绿素和类胡萝素可能对呈色有着辅助作用。2个品种叶片的解剖观察结果也体现了花青苷对叶片呈色的作用。可溶性糖是花青苷合成的原料,能一定程度上影响植物体内花青苷的含量[24-25]。本研究发现,可溶性糖含量与‘Ebony Embers’叶片中花青苷含量呈显著正相关,认为‘Ebony Embers’叶片中高含量的可溶性糖能促进叶片中花青苷的合成,最终使叶片呈现红色。
净光合速率是光合作用强弱的直观体现,反映了植物同化物生产能力的大小。本研究结果表明,虽然‘Ebony Embers’叶片的叶绿素含量比绿叶品种(‘Arapaho’)高,但一天中的Pn值始终低于‘Arapaho’,说明‘Ebony Embers’光合能力低于‘Arapaho’,而‘Ebony Embers’光合能力的降低并不是叶绿素含量的差异造成的,这与水稻(Oryza sativa)[26]及中红杨(Populus×euramericana ‘Zhanghong’)[27]的研究结果一致。2个紫薇品种净光合速率和气孔导度日变化趋势呈现出极大的一致性,说明在一定程度上气孔导度是光合作用的主要影响因素。‘Arapaho’的Pn值在午休阶段明显下降,Gs和Ci降到最低值,Ls升高到最高值,表现出较强的气孔限制。‘Ebony Embers’的Ls上午升高,影响到Pn值的下降,但其中13:00左右,Ci短暂升高,Ls短暂降低,此时限制光合作用的主要原因则由气孔限制变为非气孔限制。从2个品种光强、CO2响应特征参数可以发现,‘Arapaho’光饱和点、CO2饱和点和羧化效率显著高于‘Ebony Embers’,说明‘Arapaho’耐强光的能力较强,并且对CO2环境的适应性强,能充分利用大气中的CO2,可见相比绿叶紫薇‘Arapaho’,紫叶紫薇‘Ebony Embers’耐强光能力较低。有研究认为,花青苷含量会影响到光合色素分子对光能的吸收和利用,可能会改变光能在光合机构中的分配,从而影响光合速率[28]。另外,在桃(Prunus persica)的研究中发现,可溶性糖等光合末端产物的积累可能改变了叶片保卫细胞的渗透压进而改变了气孔导度,最终导致了净光合速率的下降[29]。本试验中,紫薇叶片净光合速率与气孔导度之间呈显著相关,‘Ebony Embers’气孔导度始终低于‘Arapaho’,而可溶性糖含量却显著高于‘Arapaho’,因此认为‘Ebony Embers’光合能力低的原因,一方面可能是叶片中高含量的花青苷影响了叶片光能的吸收,另一方面可能是叶片中较高的可溶性糖造成气孔导度下降,从而影响了净光合速率。
植物体内叶绿素荧光参数变化能反映出光合作用过程中光系统对光能的吸收、传递、耗散等反应[30]。本研究中‘Arapaho’较‘Ebony Embers’有更高的PSⅡ电子传递量子效率(ΦPSⅡ)和表观光合电子传递速率(ETR),表明‘Arapaho’的叶绿素具有较高的PSⅡ活性,能为光合作用提供更多的化学能,进一步说明绿叶紫薇较紫叶紫薇有着更高的光合能力;而‘Ebony Embers’的qN和NPQ值较‘Arapaho’高,故较‘Arapaho’有着更多的热耗散,光保护能力强于‘Arapaho’。在对紫丁香(Syringa oblata)的研究中发现,叶片中花色素苷含量与ΦPSⅡ之间呈显著负相关,与NPQ之间呈显著正相关,认为通过花色素苷吸收衰减入射光耗散过剩光能,一方面降低光化学反应的量子产额,另一方面增加了依赖于类囊体膜两侧质子梯度和叶黄素循环耗散能量的比例[31]。因此推断,‘Ebony Embers’中ΦPSⅡ和ETR的降低,非光化学淬灭的提高,可能是由于花青苷含量升高引起的。
-
表 1 群落基本特征
Table 1 Basic characteristics of the communities
样地
Sample
plot林层
Forest layer密度/
(株·hm-2)
Density/
(tree·ha-1)林分平均直径
Stand mean
DBH/cm林分平均
树高Stand
mean
height/m平均冠幅
Mean
crown
width/m蓄积量/
(m3·hm-2)
Volume/
(m3·ha-1)丰富度
RichnessShannon指数
Shannon
index1 乔木层Arbor layer 1 012 23.8 26.10 — 513.359 7 65 5.04 1 受光层Light receiving layer (LRL) 280 41.9 28.52 8.25 473.180 0 33 4.34 1 非受光层Non-light receiving layer (NLRL) 732 10.7 12.05 3.79 40.179 7 56 4.86 2 乔木层Arbor layer 1 056 22.6 24.93 — 468.140 2 53 4.59 2 受光层LRL 304 39.2 27.08 6.96 433.851 9 30 3.63 2 非受光层NLRL 752 10.0 11.53 3.36 34.288 3 47 4.41  下载: 导出CSV
下载: 导出CSV
表 2 整体关联性
Table 2 Total species association
样地
Sample plot层次
Forest layer方差比率
VRW统计量W
statistic关联性
AssociationVR偏离1
VR deviated from 11 乔木层Arbor layer 0.792 39.587 负Negative 否None significance 1 受光层LRL 0.702 35.110 负Negative 否None significance 1 非受光层NLRL 1.252 62.588 正Positive 否None significance 2 乔木层Arbor layer 0.938 46.922 负Negative 否None significance 2 受光层LRL 0.562 28.099 负Negative 是Significant 2 非受光层NLRL 0.996 49.792 负Negative 否None significance 注Notes:χ(α=0.95, f=50)2=34.764, χ(α=0.05, f=50)2=67.505。
下载: 导出CSV
表 3 1号样地各层中显著(P<0.05)和极显著(P<0.01)关联的种对
Table 3 Species pairs with significant (P < 0.05) and very significant (P < 0.01) association in sample plot 1
层次
Layer物种1
Species 1物种2
Species 2关联性
AssociationP值
P value乔木层Arbor layer 罗浮柿 木姜子 正Positive 0.009 2 乔木层Arbor layer 细枝柃 新木姜子 正Positive 0.016 9 乔木层Arbor layer 细枝柃 木荷 正Positive 0.022 6 乔木层Arbor layer 庆元冬青 新木姜子 正Positive 0.023 7 乔木层Arbor layer 香冬青 桂北木姜子 正Positive 0.029 7 乔木层Arbor layer 三花冬青 新木姜子 正Positive 0.049 9 乔木层Arbor layer 木姜子 新木姜子 正Positive 0.004 1 乔木层Arbor layer 桂北木姜子 红皮树 正Positive 0.040 0 乔木层Arbor layer 毡毛泡花树 新木姜子 正Positive 0.021 3 乔木层Arbor layer 黄瑞木 猴欢喜 负Negative 0.014 6 乔木层Arbor layer 浙江桂 野含笑 负Negative 0.046 0 乔木层Arbor layer 细齿叶柃 木荷 负Negative 0.046 4 乔木层Arbor layer 木姜子 红皮树 负Negative 0.049 7 乔木层Arbor layer 木姜子 福建山矾 负Negative 0.039 4 乔木层Arbor layer 毡毛泡花树 福建山矾 负Negative 0.028 2 乔木层Arbor layer 新木姜子 猴欢喜 负Negative 0.023 7 乔木层Arbor layer 新木姜子 光叶山矾 负Negative 0.004 0 受光层LRL 猴欢喜 香冬青 负Negative 0.046 3 受光层LRL 猴欢喜 新木姜子 负Negative 0.046 3 非受光层NLRL 米槠 福建山矾 正Positive 0.042 6 非受光层NLRL 新木姜子 细枝柃 正Positive 0.048 8 非受光层NLRL 新木姜子 庆元冬青 正Positive 0.030 4 非受光层NLRL 新木姜子 木姜子 正Positive 0.015 1 非受光层NLRL 新木姜子 笔罗子 正Positive 0.048 8 非受光层NLRL 新木姜子 红皮树 正Positive 0.048 8 非受光层NLRL 新木姜子 尖叶水丝梨 正Positive 0.041 6 非受光层NLRL 新木姜子 光叶山矾 负Negative 0.033 7 注Notes:物种名Species name:罗浮柿Diospyros morrisiana; 细枝柃Eurya loquaiana; 细齿叶柃E. nitida; 庆元冬青Ilex qingyuanensis; 香冬青I. suaveolens; 三花冬青I. triflora; 木姜子Litsea pungens; 桂北木姜子L. subcoriacea; 新木姜子Neolitsea aurata; 毡毛泡花树Meliosma rigida var. pannosa; 黄瑞木Adinandra millettii; 浙江桂Cinnamomum chekiangense; 木荷Schima superb; 红皮树Styrax suberifolius; 猴欢喜Sloanea sinensis; 野含笑Michelia skinneriana; 福建山矾Symplocos fukienensis; 光叶山矾S.lancifolia; 米槠Castanopsis carlesii; 笔罗子Meliosma rigida; 尖叶水丝梨Sycopsis dunnii.
下载: 导出CSV
表 4 2号样地各层中显著(P<0.05)和极显著(P<0.01)关联的种对
Table 4 Species pairs with significant (P < 0.05) and very significant (P < 0.01) association in sample plot 2
层次
Layer物种1
Species 1物种2
Species 2关联性
AssociationP值
P value乔木层Arbor layer 米槠 乐东拟单性
木兰正Positive 0.030 42 乔木层Arbor layer 拉氏栲 山黄皮 正Positive 0.031 76 乔木层Arbor layer 山杜英 木姜子 正Positive 0.026 55 乔木层Arbor layer 三花冬青 木姜子 正Positive 0.047 70 乔木层Arbor layer 毡毛泡花树 野含笑 正Positive 0.048 26 乔木层Arbor layer 浙江桂 香冬青 负Negative 0.048 81 乔木层Arbor layer 山杜英 光叶山矾 负Negative 0.024 83 乔木层Arbor layer 庆元冬青 木姜子 负Negative 0.043 84 乔木层Arbor layer 木荷 猴欢喜 负Negative 0.018 62 乔木层Arbor layer 猴欢喜 光叶山矾 负Negative 0.024 83 受光层LRL 浙江桂 猴欢喜 负Negative 0.025 99 非受光层NLRL 山杜英 弯蒴杜鹃 正Positive 0.034 69 非受光层NLRL 山杜英 光叶山矾 负Negative 0.005 18 非受光层NLRL 猴欢喜 光叶山矾 负Negative 0.034 54 注Notes:物种名Species name:米槠Castanopsis carlesii; 拉氏栲C. lamontii; 山杜英Elaeocarpus sylvestris; 香冬青Ilex suaveolens; 三花冬青I. triflora; 庆元冬青I. qingyuanensis; 毡毛泡花树Meliosma rigida var. pannosa; 浙江桂Cinnamomum chekiangense; 乐东拟单性木兰Parakmeria lotungensis; 山黄皮Randia cochinchinensis; 木姜子Litsea pungens; 野含笑Michelia skinneriana; 光叶山矾Symplocos lancifolia; 木荷Schima superb; 猴欢喜Sloanea sinensis; 弯蒴杜鹃Rhododendron henryi.
下载: 导出CSV
-
[1] CALLAWAY R M, WALKER L R. Competition and facilitation: a synthetic approach to interactions in plant communities[J]. Ecology, 1997, 78(7): 1958-1965. doi: 10.1890/0012-9658(1997)078[1958:CAFASA]2.0.CO;2
[2] 彭少麟, 周厚诚, 郭少聪, 等.鼎湖山地带性植被种间联结变化研究[J].植物学报, 1999, 41(11): 1239-1244. doi: 10.3321/j.issn:1672-9072.1999.11.020 PENG S L, ZHOU H C, GUO S C, et al. Studies on the changes in interspecific association of zonal vegetation in Dinghushan[J]. Acta Botanica Sinica, 1999, 41(11): 1239-1244. doi: 10.3321/j.issn:1672-9072.1999.11.020
[3] CALLAWAY R M, BROOKER R W, CHOLER P, et al. Positive interactions among alpine plants increase with stress[J]. Nature, 2002, 417: 844-848. doi: 10.1038/nature00812
[4] CHAMBERLAIN S A, BRONSTEIN J L, RUDGERS J A. How context dependent are species interactions?[J]. Ecology Letters, 2014, 17(7): 881-890. doi: 10.1111/ele.12279
[5] PATERNO G B, SIQUEIRA FILHO J A, GANADE G. Species-specific facilitation, ontogenetic shifts and consequences for plant community succession[J]. Journal of Vegetation Science, 2016, 27(3): 606-615. doi: 10.1111/jvs.12382
[6] MORIN P J. Community ecology[M]. 2nd ed. Oxford: Wiley-Blackwell, 2011: 29-67, 185, 205.
[7] 孙祯元, 郜超, 贾艳梅, 等.榆林沙地植物种间联结及共生利害辨析[J].北京林业大学学报, 2015, 37(11):41-47. doi: 10.13332/j.1000-1522.20150002 SUN Z Y, GAO C, JIA Y M, et al. Analysis of mutual benefit and damage of interspecific association of plant species in sandy area in Yulin, Shaanxi of western China[J]. Journal of Beijing Forestry University, 2015, 37(11):41-47. doi: 10.13332/j.1000-1522.20150002
[8] 洪伟, 陈鸣煊.闽北主要阔叶树种种间联结测定及其应用[J].林业科学, 1990, 26(3): 175-181. http://www.cnki.com.cn/Article/CJFDTotal-LYKE199002010.htm HONG W, CHEN M X. Calculation and application of interspecific association of broad-leaved trees in the north Fujian[J]. Scientia Silvae Sinicae, 1990, 26(3): 175-181. http://www.cnki.com.cn/Article/CJFDTotal-LYKE199002010.htm
[9] BERGOTTINI V M, HERVÉ V, SOSA D A, et al.Exploring the diversity of the root-associated microbiome of Ilex paraguariensis St. Hil.(Yerba Mate)[J]. Applied Soil Ecology, 2017, 109: 23-31. doi: 10.1016/j.apsoil.2016.09.013
[10] 罗梅, 郑小贤.福建将乐林场常绿阔叶次生林主要种群种间联结性研究[J].西北农林科技大学学报(自然科学版), 2016, 44(8): 135-142. http://d.old.wanfangdata.com.cn/Periodical/xbnydxxb201608020 LUO M, ZHENG X X. Inter-specific correlation of main populations in evergreen broad-leaved secondary forest in Jiangle Forest Farm[J]. Journal of Northwest A&F University (Natural Science Edition), 2016, 44(8): 135-142. http://d.old.wanfangdata.com.cn/Periodical/xbnydxxb201608020
[11] 胡理乐, 江明喜, 党海山, 等.从种间联结分析濒危植物毛柄小勾儿茶在群落中的地位[J].植物生态学报, 2005, 29(2): 258-265. doi: 10.3321/j.issn:1005-264X.2005.02.011 HU L L, JIANG M X, DANG H S, et al. Community studies on the status of the endangered plant Berchemiella wilsonii var. pubipetiolata, using interspecific association analysis[J]. Acta Phytoecologica Sinica, 2005, 29(2): 258-265. doi: 10.3321/j.issn:1005-264X.2005.02.011
[12] WALKER L R, VITOUSEK P M.An invader alters germination and growth of a native dominant tree in Hawaii[J]. Ecology, 1991, 72(4): 1449-1455. doi: 10.2307/1941117
[13] KUEBBING S E, NUÑEZ M A. Negative, neutral, and positive interactions among nonnative plants: patterns, processes, and management implications[J]. Global Change Biology, 2015, 21(2):926-934. doi: 10.1111/gcb.12711
[14] 钟章成.常绿阔叶林生态学研究[M].重庆:西南师范大学出版社, 1988: 4-7. ZHONG Z C. Ecology study on evergreen broadleaved forest[M]. Chongqing: Southwest China Normal University Press, 1988: 4-7.
[15] 贺金生, 陈伟烈, 李凌浩.中国中亚热带东部常绿阔叶林主要类型的群落多样性特征[J].植物生态学报, 1998, 22(4): 303-311. doi: 10.3321/j.issn:1005-264X.1998.04.003 HE J S, CHEN W L, LI L H. Community diversity of the main types of the evergreen broad-leaved forest in the eastern part of the middle subtropical China[J]. Acta Phytoecologica Sinica, 1998, 22(4): 303-311. doi: 10.3321/j.issn:1005-264X.1998.04.003
[16] 王伯荪, 彭少麟.鼎湖山森林群落分析Ⅱ:物种联结性[J].中山大学学报(自然科学版), 1983(4): 29-35. doi: 10.1002-ajmg.a.33115/ WANG B S, PENG S L. Analysis on the forest commumities of Dinghushan (Ⅱ): species association[J]. Acta Scientiarum Naturalium Universitatis Sunyatseni, 1983(4): 29-35. doi: 10.1002-ajmg.a.33115/
[17] 谢亮秀, 游水生, 吴启銮.米槠群落优势种群间联结性研究[J].内蒙古林学院学报(自然科学版), 1999, 21(2): 31-34. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=QK199900501290 XIE L X, YOU S S, WU Q L. Research of interspecific association on the dorminant 15 populations of Castanopsis carlesii community in Fujian Wupin[J]. Journal of Inner Mongolia Forestry College, 1999, 21(2): 31-34. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=QK199900501290
[18] 黄云鹏.武夷山米槠林主要树种种间关联性[J].山地学报, 2008, 26(6): 692-698. doi: 10.3969/j.issn.1008-2786.2008.06.007 HUANG Y P. Studies on the relationship of dominant species in the Castanopsis carlesii forest in Wuyishan Scenery District[J]. Journal of Mountain Science, 2008, 26(6): 692-698. doi: 10.3969/j.issn.1008-2786.2008.06.007
[19] 金俊彦, 覃文更, 谭卫宁, 等.濒危植物单性木兰群落主要种群种间联结性研究[J].西部林业科学, 2013, 42(3): 86-94. doi: 10.3969/j.issn.1672-8246.2013.03.015 JIN J Y, QIN W G, TAN W N, et al. Interspecific association among dominant populations in endangered plant Kmeria septentrionalis communities[J]. Journal of West China Forestry Science, 2013, 42(3): 86-94. doi: 10.3969/j.issn.1672-8246.2013.03.015
[20] 中国植被编委会.中国植被[M].北京:科学出版社, 1980: 279-306. Vegetation of China Editorial Committee. Vegetation of China[M]. Beijing: Science Press, 1980: 279-306.
[21] 陈灵芝, 陈清朗, 刘文华.中国森林多样性及其地理分布[M].北京:科学出版社, 1997: 132-159. CHEN L Z, CHEN Q L, LIU W H. Forest diversity and its geographical distribution in China[M]. Beijing: Science Press, 1997: 132-159.
[22] 黄清麟, 李元红.中亚热带天然阔叶林可持续经营的若干问题[J].福建林学院学报, 2000, 20(1): 1-4. doi: 10.3969/j.issn.1001-389X.2000.01.001 HUANG Q L, LI Y H. Several problems about sustainable management of natural broad-leaved forest in the mid-subtropical zone[J]. Journal of Fujian College of Forestry, 2000, 20(1): 1-4. doi: 10.3969/j.issn.1001-389X.2000.01.001
[23] VEECH J A. A probabilistic model for analysing species co-occurrence[J]. Global Ecology and Biogeography, 2013, 22(2): 252-260. doi: 10.1111/j.1466-8238.2012.00789.x
[24] 何友钊.建瓯县万木林保护区史事考[M]//中国林学会林业史学会.林史文集(第一辑).北京: 中国林业出版社, 1990. HE Y Z. Examining for the historical events in Wanmulin Nature Reserve[M]//Society of Forestry Historiography of Chinese Society of Forestry. Corpus of Forestry Historiography Ⅰ. Beijing: China Forestry Publishing House, 1990.
[25] 黄清麟, 李志明, 郑群瑞.福建中亚热带天然阔叶林理想结构探讨[J].山地学报, 2003, 21(1): 116-120. doi: 10.3969/j.issn.1008-2786.2003.01.019 HUANG Q L, LI Z M, ZHENG Q R. A discussion on the ideal structure of natural broad-leaved forest in the mid-subtropical zong of Fujian Province[J]. Journal of Mountain Science, 2003, 21(1): 116-120. doi: 10.3969/j.issn.1008-2786.2003.01.019
[26] 庄崇洋, 黄清麟, 马志波, 等.中亚热带天然阔叶林林层划分新方法:最大受光面法[J].林业科学, 2017, 53(3): 1-11. http://d.old.wanfangdata.com.cn/Periodical/lykx201703001 ZHUANG C Y, HUANG Q L, MA Z B, et al. A new method of storey identification of natural broad-leaved forests in mid-subtropical zone: maximum light receiving plane[J].Scientia Silvae Sinicae, 2017, 53(3): 1-11. http://d.old.wanfangdata.com.cn/Periodical/lykx201703001
[27] SCHULTER D. A variance test for detecting species associations with some example applications[J]. Ecology, 1984, 65(3): 998-1005. doi: 10.2307/1938071
[28] GRIFFITH D M, VEECH J A, MARSH C J. Cooccur: probabilistic species co-occurrence analysis in R[J]. Journal of Statistical Software, 2016, 69(2): 1-17. http://cn.bing.com/academic/profile?id=a81663adcffe8edbcd4bb4abb9ea7573&encoded=0&v=paper_preview&mkt=zh-cn
[29] ASCHEHOUG E T, BROOKER R, ATWATER D Z, et al. The mechanisms and consequences of interspecific competition among plants[J]. Annual Review of Ecology, Evolution, and Systematics, 2016, 47: 263-281. doi: 10.1146/annurev-ecolsys-121415-032123
[30] 马克平.监测是评估生物多样性保护进展的有效途径[J].生物多样性, 2011, 19(2):125-126. http://d.old.wanfangdata.com.cn/Periodical/swdyx201102001 MA K P. Assessing progress of biodiversity conservation with monitoring approach[J]. Biodiversity Science, 2011, 19(2):125-126. http://d.old.wanfangdata.com.cn/Periodical/swdyx201102001
[31] 宋永昌, 阎恩荣, 宋坤.中国常绿阔叶林8大动态监测样地植被的综合比较[J].生物多样性, 2015, 23(2): 139-148. http://d.old.wanfangdata.com.cn/Periodical/swdyx201502002 SONG Y C, YAN E R, SONG K. Synthetic comparison of eight dynamics plots in evergreen broadleaf forests, China[J]. Biodiversity Science, 2015, 23(2): 139-148. http://d.old.wanfangdata.com.cn/Periodical/swdyx201502002
[32] BRUELHEIDE H, BÖHNKE M, BOTH S, et al. Community assembly during secondary forest succession in a Chinese subtropical forest[J]. Ecological Monographs, 2011, 81(1): 25-41. doi: 10.1890/09-2172.1
[33] BOTH S, FANG T, BÖHNKE M, et al. Lack of tree layer control on herb layer characteristics in a subtropical forest, China[J]. Journal of Vegetation Science, 2011, 22(6): 1120-1131. doi: 10.1111/j.1654-1103.2011.01324.x
[34] WIEGAND T, HUTH A, GETZIN S, et al. Testing the independent species' arrangement assertion made by theories of stochastic geometry of biodiversity[J]. Proceedings of the Royal Society (Biological Sciences), 2012, 279: 3312-3320. doi: 10.1098/rspb.2012.0376
[35] WANG X G, WIEGAND T, KRAFT N J B, et al. Stochastic dilution effects weaken deterministic effects of niche-based processes in species rich forests[J]. Ecology, 2016, 97(2):347-360. doi: 10.1890/14-2357.1
[36] HUBBELL S P. Neutral theory and the evolution of ecological equivalence[J]. Ecology, 2006, 87(6): 1387-1398. doi: 10.1890/0012-9658(2006)87[1387:NTATEO]2.0.CO;2
[37] PUNCHI-MANAGE R, WIEGAND T, WIEGAND K, et al. Neighborhood diversity of large trees shows independent species patterns in a mixed dipterocarp forest in Sri Lanka[J]. Ecology, 2015, 96(7): 1823-1834 doi: 10.1890/14-1477.1
-
期刊类型引用(21)
1. 张蔓蔓,郑聪慧,徐振华,李新利,杜克久,田芸,孟李冰,柳明妹,聂佳佳,邢争. 2种红果臭椿表型特征及色素组分相关性. 东北林业大学学报. 2025(01): 23-34+42 .  百度学术
百度学术
2. 刘玉军,秦波,李媚,黄耀恒,刘芳,黄欣,刘莉. 黑紫色叶紫薇品种‘赤红’叶片转录组测序及注释分析. 湖南农业科学. 2024(01): 1-6 . 百度学术
3. 胡平. 光照条件对不同类型绿地南天竹叶色变化的影响. 安徽林业科技. 2024(02): 24-28 . 百度学术
4. Lu Feng,Ping Shen,Xiufeng Chi,Yang Zhou,Jieru Liu,Tangren Cheng,Jia Wang,Qixiang Zhang,Ming Cai,Huitang Pan. The anthocyanin formation of purple leaf is associated with the activation of LfiHY5 and LfiMYB75 in crape myrtle. Horticultural Plant Journal. 2024(05): 1230-1246 . 必应学术
5. 崔祺,吴昀,李东泽,吴凡,韩蕊莲,黄均华,胡绍庆. 彩叶桂叶片发育过程中叶色表型与色素成分变化. 南京林业大学学报(自然科学版). 2023(02): 79-86 . 百度学术
6. 李茂娟,崔强,徐毓泽,张海霞,廖祯妮,何才生. 紫叶紫薇高接换冠技术研究. 中国农学通报. 2023(25): 21-26 . 百度学术
7. 刘雄盛,尹国平,肖玉菲,蒋燚,王仁杰,黄荣林,姜英,王勇. 枫香叶片变色期全长转录组测序及分析. 广西植物. 2023(09): 1710-1720 . 百度学术
8. 郑聪慧,徐振华,王玉忠,张蔓蔓,杜克久,李向军,刘春鹏. 四种臭椿不同叶位叶片色素组成及分布变化. 中国农学通报. 2023(32): 56-65 . 百度学术
9. 尹国平,刘雄盛,蒋燚,王勇,杨继生,肖玉菲,黄荣林,姜英. 枫香变红过程中叶片组织结构、光合特性及色素含量变化研究. 广西植物. 2022(07): 1213-1221 . 百度学术
10. 邵雯雯,何钢,乔中全,曾慧杰,蔡能,石炳霖. 6个紫薇品种叶片色彩变化及其与色素含量的相关性. 西北林学院学报. 2022(05): 104-110+123 . 百度学术
11. 张阳,钱多,马喆. 彩叶树种叶色影响因素及光合特性研究进展. 安徽农业科学. 2022(20): 12-17 . 百度学术
12. 王永涛,杨宁,李贵学,于林倩,姜锡川. 彩叶植物SPAD值及其荧光参数的异质性. 绿色科技. 2022(23): 92-97 . 百度学术
13. 马赞留. 彩叶紫薇育种及繁育技术研究进展. 农业开发与装备. 2022(12): 238-240 . 百度学术
14. 吴焦焦,田秋玲,谭星,乐佳兴,张文,高岚,李林珂,王一诺,刘芸. 氮磷钾肥配施对黄栌生长和叶片呈色的影响. 林业科学. 2021(11): 179-189 . 百度学术
15. 卢玉生,官凤英,彭超,尹子旭,张美曼,詹美春,晏颖杰. 竹笋截梢对绿竹生长及叶绿素荧光特性的影响. 浙江农林大学学报. 2020(01): 51-59 . 百度学术
16. 陈露,李家明,周相助,许茹,林义章,钟凤林. 外施苹果酸对紫色小白菜花青苷合成关键基因的影响. 农业生物技术学报. 2020(03): 407-419 . 百度学术
17. 肖杰,薛欢,苑景淇,于忠亮,李成宏,王梅芳,周梅妹,杜凤国. 3个紫薇品种幼苗光响应特性. 北华大学学报(自然科学版). 2020(03): 311-319 . 百度学术
18. 崔舜,邱国金,吴茜,戴静,徐炅,夏爱华. 彩叶紫薇新品种红火球与仑山1号的叶色及生理变化特性. 贵州农业科学. 2020(09): 16-21 . 百度学术
19. 肖杰,薛欢,苑景淇,于忠亮,李成宏,王梅芳,周梅妹,杜凤国. 3个紫薇品种实生苗和扦插苗光合特性比较及光合效率评价. 西部林业科学. 2020(06): 85-93 . 百度学术
20. 吕运舟,董筱昀,杨小鑫,孙海楠,何开跃,黄利斌. 黄山栾树新品种‘金焰彩栾’叶片呈色的生理特性及影响因子分析. 植物资源与环境学报. 2020(06): 51-56 . 百度学术
21. 宋鹏,丁彦芬,卓启苗,李涵,王亚楠,徐子涵,蔡慧. 3种卫矛属植物叶片秋冬季转色期生理生化特征研究. 西北植物学报. 2019(04): 669-676 . 百度学术
其他类型引用(10)



计量
- 文章访问数: 1538
- HTML全文浏览量: 460
- PDF下载量: 31
- 被引次数: 31
