
Vegetation phenology dynamic and its responses to meteorological factor changes in the Mu Us Desert of northern China
-
摘要:目的监测沙区区域植被物候,并确定植被物候与气候要素的关系,对理解沙区生态系统对气候变化的动态响应至关重要。方法本研究利用1982年1月至2015年12月的归一化差值植被指数(NDVI)时间序列数据,通过高阶曲线拟合的方法提取毛乌素沙地植被物候,包括生长季开始日期、结束日期和长度,分析其时间变化和空间特征,并利用偏相关性分析法确定其与气象因子(气温和降水量)的关系。结果毛乌素沙地植被生长季开始日期集中在第96~144天,生长季结束日期集中于第280~300天,生长季长度介于135~195d。从东到西生长季开始日期逐渐推迟,生长季长度逐渐缩短,生长季结束日期没有明显变化。在过去34年中,毛乌素沙地植被生长季开始日期和长度分别呈现显著提前和延长的趋势,分别为0.54和0.94d/a,生长季结束日期呈现推迟趋势,但该趋势并不显著。毛乌素沙地植被的生长季开始日期主要受2—4月的平均气温影响,结束日期主要受8—10月的累计降水量的影响。结论毛乌素沙地植被物候特征发生了明显变化,主要体现在生长季的提前和延长,这对该区域植被恢复与荒漠化的逆转具有一定的促进作用。Abstract:ObjectiveMonitoring regional vegetation phenology in sand area and determining the relations between vegetation phenology and meteorological factors are crucial to understand the dynamic response of sand area ecosystem to climate change.MethodIn this study, through normalized difference vegetation index (NDVI) time series data, we extracted vegetation phenology by Polyfit-Maximum method, and analyzed the trends and relationships between vegetation phenology and climatic factors (air temperature and precipitation) in the Mu Us Desert of northern China over the past three decades (1982-2015).ResultIn the Mu Us Desert, The beginning date of growing season concentrated in the 96th-144th day, the ending date of growing season concentrated in the 280th-300th day, and the length of growing season ranged in 135-195 days. From east to west, the beginning date of growing season postponed gradually and growing season length shortened gradually, while there was no obvious change in the ending date of growing season. In the past 34 years, the beginning date and length of growing season for vegetation in the Mu Us Desert showed significantly advanced and prolonged trend, and the changing rates were 0.54 day/year and 0.94 day/year, respectively. While the ending date of growing season showed postponed trend but not delayed obviously. The beginning date of growing season for vegetation in the Mu Us Desert was mainly affected by the mean air temperature from February to April, and its ending date was mainly affected by the accumulated precipitation from August to October.ConclusionThe vegetation phenology characteristics have changed significantly in Mu Us Desert, which was mainly reflected in the advancement and extension of growing season. This will promote the vegetation restoration and desertification reversion in the study area.
-
Keywords:
- NDVI /
- vegetation phenology /
- Mu Us Desert /
- meteorological factor /
- spatiotemporal dynamics
-
油橄榄(Olea europaea)主要分布于地中海地区,是一种重要的木本油料作物。橄榄油中富含健康的不饱和脂肪酸、酚类化合物和维生素E等营养物质,不仅是地中海式饮食的重要成分,而且远销美国、墨西哥、巴西、印度、澳大利亚等国家[1]。近年来,全球橄榄油需求量增加,产量却十分有限。要持续扩大油橄榄的适生区,增加橄榄油的产量,种质选育创制尤为关键。随着测序技术的飞速发展,油橄榄已完成了高质量基因组文件的组装和注释[2],并获得了丰富的转录组数据,这为我们借助分子育种选育抗逆性强、产油率高的油橄榄奠定了基础。
过氧化物酶(peroxidase,PRX)是一类广泛存在于生物体内的酶,它通过将过氧化氢还原为水来催化底物的氧化。根据其蛋白序列和结构特征,PRX可分为2类,血红素类和非血红素类;血红素PRX又可进一步分为动物型和非动物型;非动物的PRX包含3个小类,Ⅰ、Ⅱ和Ⅲ类,这3小类PRX具有非常相似的三维结构,但氨基酸序列同源性较低,且具有不同的功能和反应机制[3]。Ⅰ类PRX广泛分布于非动物性生命体内,其在原核生物中的分布表明其他2类PRX可能起源于Ⅰ类PRXs[4],Ⅰ类PRX包含抗坏血酸PRX、细胞色素c PRX和过氧化氢酶,主要功能是消除过量的H2O2[5-6]。Ⅱ类PRX仅分布于真菌中,包含锰PRX、木质素PRX和其他PRX[7],主要功能是降解木质素。Ⅲ类PRX分布于陆生植物体内,是一个庞大的多基因家族,通常分泌进入细胞壁或其周围基质或液泡中。Ⅲ类PRX广泛参与植物生命周期内的一系列的生理活动[8],比如细胞壁形成[9]、伤口愈合[10]、生长素代谢[11-12]等。另外,Ⅲ类PRX通过调节多种活性氧水平响应生物胁迫和非生物胁迫[12],因此,研究Ⅲ类PRX基因功能对作物遗传育种具有十分重要的作用。
目前,拟南芥(Arabidopsis thaliana)[13]、水稻(Oryza sativa)[14]、玉米(Zea mays)[15]、毛果杨(Populus trichocarpa)[16]、白梨(Pyrus bretschneideri)[17]、葡萄(Vitis vinifera)[18]、木薯(Manihot esculenta)[19]和小麦(Triticum aestivum)[20]等植物的Ⅲ类PRX基因全基因组鉴定和分析已经完成,这些物种中有47 ~ 138个不等的Ⅲ类PRX基因。单个Ⅲ类PRX基因功能的研究也日益丰富,例如,AtPRX4、AtPRX52和AtPRX72拟南芥突变体木质素含量降低,木质素组成发生改变[21-23]。烟草(Nicotiana tabacum)中过表达AtPRX64促进根系生长,减少活性氧和铝在根系中的积累,从而提高对铝胁迫的耐受性[24];拟南芥中过表达OsPRX38促进胼胝质沉积、抗氧化酶活性和叶绿素含量上升,从而提高植物对砷胁迫的耐受性[25]。然而,关于油橄榄Ⅲ类PRX基因家族的全基因组鉴定及基因功能研究仍未见报道。本研究利用最近组装注释的油橄榄基因组和公共数据库中的转录组信息,完成了油橄榄Ⅲ类PRX家族的鉴定、进化和表达分析。研究发现:油橄榄是目前已知Ⅲ类PRX家族成员最多的双子叶植物,其复制基因的亚功能化和新功能化对油橄榄在地中海地区的广泛分布具有十分重要的意义。
1. 材料与方法
1.1 油橄榄Ⅲ类PRX基因家族成员的鉴定
油橄榄基因组文件下载于NGDC(https://bigd.big.ac.cn/search/?dbId=gwh&q=Olea%20europaea&page=1),Ⅲ类PRX基因家族的保守结构域(PF00141)的HMM模型下载于pfam(http://pfam.xfam.org/family/PF00141#tabview=tab6)。利用PF00141的HMM模型,在油橄榄蛋白质注释文件(GWHAOPM00000000.Protein.faa)中进行HMM模型搜索(hmmsearch),得到120条蛋白序列。利用拟南芥的73条PRX蛋白序列[13]在油橄榄基因组文件(GWHAOPM00000000.Protein.faa)中进行本地化搜索(blastp),E值设置为10−6,得到119条序列。2种方法得到的共有序列为119条。将119条序列上传至Pfam(http://pfam.xfam.org/search#tabview=tab1)和NCBI-CDD(https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)数据库中,预测其是否含有Ⅲ类PRX基因家族的保守结构域从而进行筛选。Pfam鉴定结果表明,119条序列均含有PF00141结构域;NCBI-CDD鉴定结果表明,13条序列含有不完整的分泌类过氧化物酶保守结构域(secretory_peroxidase,cd00693)。因此,最后选定106条序列作为油橄榄Ⅲ类PRX基因成员。利用Prot-Param(https://web.expasy.org/compute_pi/)预测等电点和分子量;利用SignalP-5.0 Server(http://www.cbs.dtu.dk/services/SignalP/)预测信号肽Sec/SPI的存在概率及长度,Signal peptide likelihood值大于0.5的序列视为含有信号肽,反之认为不含有信号肽;利用Wolfpsort(https://wolfpsort.hgc.jp/)预测亚细胞定位。
1.2 PRX蛋白系统进化树的构建
利用TBtools v1.095中的Fasta Extract从油橄榄基因组文件中提取得到106个OePRX的蛋白序列,从前人的论文中下载得到拟南芥和毛果杨PRX蛋白序列[16],其中,拟南芥73条,毛果杨93条。利用MEGA7.0中的ClustalW比对272条全长的PRX蛋白序列,并使用邻接法构建系统进化树,设置替代模型为Poisson model,设置缺失数据处理方式为Pairwise deletion,设置bootstrap值为1 000。
1.3 OePRX染色体定位和复制基因分析
利用TBtools v1.095中的Fasta Stats从油橄榄基因组文件中获取23条染色体的长度和OePRX基因家族成员的位置信息,利用MapChart绘制OePRX基因家族在23条染色体上的分布,需注意的是,OePRX基因家族成员中100个基因分布于染色体上,但6个基因分布于Contig上,这6个基因未在图中展示。利用TBtools v1.095中的One Step MCScanX分析OePRX基因家族中的复制基因;利用MEGA7.0中的Muscle (Codons)比对复制基因对的编码区(CDS),再利用KaKs_Calculator2.0计算复制基因对的非同义替换(Ka)和同义替换(Ks)值,设置计算方式为YN。拟南芥基因组文件下载自http://ensembl.gramene.org/Arabidopsis_thaliana/Info/Index,毛果杨基因组文件下载自https://phytozome.jgi.doe.gov。利用同样的方法分析油橄榄和拟南芥及毛果杨PRX之间的复制基因,并计算Ka和Ks值。利用TBtools v1.095中的Advanced Circos绘制油橄榄PRX基因线性图;用Multiple Synteny Plot绘制油橄榄与拟南芥和毛果杨PRX基因的线性图。
1.4 OePRX基因序列分析
利用TBtools v1.095中的Fasta Extract从油橄榄基因组文件提取OePRX基因家族成员的DNA序列和CDS序列,再利用GSDS2.0(http://gsds.gao-lab.org/)绘制其基因结构图。利用在线网站MEME(https://meme-suite.org)预测OePRX基因的保守基序,预测基序的个数设置为10,其他参数默认。利用TBtools v1.095中的Fasta Extract从油橄榄基因组文件中获取106个OePRX基因上游1 500 bp的序列作为启动子,并将其提交至PlantCARE(http://bioinformatics.psb.ugent.be)预测启动子上的顺式作用元件,最后对结果进行分析整理。
1.5 OePRX基因表达模式分析
利用NCBI(https://www.ncbi.nlm.nih.gov/)中油橄榄不同组织[26](根、茎、叶、花、果和分生组织)、油橄榄枝条响应干热胁迫及复水[27]和油橄榄叶片响应水涝胁迫的RNA-seq数据(未发表)分析OePRX基因家族的表达模式。使用sra explorer获取测序数据的FTP地址,使用fastp去除接头和低质量的reads,利用hisat2比对并使用htseq-count获取基因的原始的读长数目(reads counts),使用DEseq2进行差异表达分析。自行编写R脚本将原始读长数目转换为tpm(每千个碱基的转录每百万映射读取的转录本,均一化后的read counts值)。差异基因鉴定标准为FDR ≤ 0.05,并且|log2(Fold Change)| ≥ 2(Fold Change为差异倍数)。取log2(tpm + 1),利用R语言绘制热图。
1.6 实时荧光定量PCR
油橄榄品种“科罗莱卡”种植于中国林科院林木遗传育种国家重点实验室温室,选择3株长势均一的3年生油橄榄幼苗,剪取叶片、茎、根和果实,于液氮中带回实验室后冻存于−80 ℃冰箱内。利用MiniBEST植物RNA提取试剂盒(TaKaRa,9769)提取不同组织的总RNA;利用PrimeScript™ RT Master Mix(Takara,RR036A)反转录合成cDNA;利用Primer Premier5.0设计引物;利用TB Green® Premix Ex Taq™ Ⅱ(Tli RNaseH Plus)(Takara,RR820A)在Roche Light Cycler 480上进行实时荧光定量PCR(qRT-PCR),反应体系如下:总体系20 μL,包括TB Green Premix Ex Taq Ⅱ(Tli RNaseH Plus)(2×)10 μL,cDNA 2 μL,正、反向引物各0.8 μL,去离子水6.4 μL。反应程序为2步法,详见试剂盒说明书。以Actin[28]为内参基因,利用2−△△CT法计算基因相对表达量。
2. 结果与分析
2.1 OePRX基因家族成员的鉴定
血红素过氧化物酶(haem peroxidase)的保守结构域是PF00141,这类酶大都含有2个α结构域,中间嵌有血红素基团[29]。利用PF00141的HMM模型和73条AtPRX蛋白在油橄榄蛋白质序列文件中分别进行hmmsearch和blastp,将2种方法得到的结果取交集,得到了119条候选PRX。植物Ⅲ类过氧化物酶特有的结构域是分泌类过氧化物酶保守结构域(secretory_peroxidase,cd00693),这类酶含有4个保守的二硫桥和2个保守的钙离子结合位点[30]。根据NCBI-CDD预测结果,删除分泌类过氧化物酶保守结构域不完整的13条序列,最后得到了106条油橄榄Ⅲ类PRX蛋白序列(表1)。根据其在染色体上的分布情况,将其命名为OePRX1-OePRX106。OePRX蛋白的氨基酸个数最小为240,最大为509;分子量最小为26.55 kDa,最大为58.00 kDa;等电点变化范围为4.45 ~ 9.75,其中74个OePRX蛋白(69.8%)大于等于7。通过在线预测发现:75个OePRX蛋白(70.8%)含有Sec/SPI信号肽,信号肽长度变化范围为19 ~ 32个氨基酸;值得注意的是,组14中的10个OePRX蛋白均不含有信号肽。亚细胞定位预测结果显示:含有信号肽的OePRX蛋白大多位于叶绿体、质膜、液泡膜和内质网(50个OePRX蛋白,占含有信号肽的OePRX蛋白总数的66.7%),也有一部分位于细胞外和细胞质(25个OeORXs蛋白,占含有信号肽的OePRX总数的33.3%);不含信号肽的OePRX蛋白(31个OePRX蛋白)中,11个OePRX蛋白位于细胞外和细胞质,其余位于叶绿体、内质网、细胞核和质膜。
表 1 油橄榄Ⅲ类PRX 基因的命名及其理化性质Table 1. Nomenclature and physicochemical properties of class Ⅲ PRX genes from olive名称 Name ID 内含子相位
Intron phase蛋白质长度
Protein length/aa分子量
Molecular mass/kDa等电点
Isoelectric point信号肽
Signal peptide预测的亚细胞定位
Predicted subcellular localizationOePRX1 EVM0022477 0 344 38.40 6.10 无 No 胞外 Extracellular OePRX2 EVM0032926 0 266 29.38 8.97 无 No 细胞质 Cytosol OePRX3 EVM0020804 0 327 35.65 9.21 有 Yes 叶绿体 Chloroplast OePRX4 EVM0047797 100 327 35.92 8.24 无 No 细胞核 Nucleus OePRX5 EVM0009292 100 316 34.40 8.72 有 Yes 液泡膜 Vacular membrane OePRX6 EVM0060238 0 330 35.97 8.89 无 No 叶绿体 Chloroplast OePRX7 EVM0022620 12001000 276 30.39 8.66 无 No 细胞质 Cytosol OePRX8 EVM0017319 1200201 279 30.58 7.70 无 No 细胞质 Cytosol OePRX9 EVM0047458 20010 250 27.43 5.67 无 No 细胞质 Cytosol OePRX10 EVM0057886 0 319 34.48 6.98 有 Yes 叶绿体 Chloroplast OePRX11 EVM0047784 0 339 37.00 7.48 有 Yes 质膜 Plasma membrane OePRX12 EVM0039807 20 358 38.84 7.51 有 Yes 叶绿体 Chloroplast OePRX13 EVM0011450 0 317 34.35 9.14 有 Yes 叶绿体 Chloroplast OePRX14 EVM0019223 1 309 35.27 9.41 有 Yes 胞外 Extracellular OePRX15 EVM0012728 1 320 36.91 9.20 有 Yes 细胞质 Cytosol OePRX16 EVM0000231 0 336 37.28 7.08 有 Yes 叶绿体 Chloroplast OePRX17 EVM0024793 0 322 35.21 8.38 有 Yes 叶绿体 Chloroplast OePRX18 EVM0042519 0 321 34.96 8.76 有 Yes 叶绿体 Chloroplast OePRX19 EVM0059982 0 321 34.90 8.77 有 Yes 叶绿体 Chloroplast OePRX20 EVM0019685 0 333 36.73 5.68 无 No 质膜 Plasma membrane OePRX21 EVM0058087 0 324 35.74 8.53 有 Yes 细胞质 Cytosol OePRX22 EVM0027348 0 323 35.02 5.82 有 Yes 液液泡膜 Vacular membrane OePRX23 EVM0060830 0 331 36.34 9.41 有 Yes 胞外 Extracellular OePRX24 EVM0015621 0 247 27.39 8.88 无 No 细胞核 Nucleus OePRX25 EVM0010445 12101221 327 35.56 9.26 无 No 叶绿体 Chloroplast OePRX26 EVM0006524 0 338 37.53 6.45 有 Yes 胞外 Extracellular OePRX27 EVM0042768 22 304 32.63 6.58 有 Yes 叶绿体 Chloroplast OePRX28 EVM0048359 0 355 37.59 8.35 有 Yes 叶绿体 Chloroplast OePRX29 EVM0028664 0 317 34.49 9.09 有 Yes 液泡膜 Vacular membrane OePRX30 EVM0035621 0 327 34.85 5.51 有 Yes 胞外 Extracellular OePRX31 EVM0048816 100 324 36.67 6.26 有 Yes 胞外 Extracellular OePRX32 EVM0003637 1002 350 39.07 8.83 有 Yes 胞外 Extracellular OePRX33 EVM0019330 0 333 36.44 9.48 有 Yes 液泡膜 Vacular membrane OePRX34 EVM0033281 2100102 273 30.15 7.73 无 No 细胞质 Cytosol OePRX35 EVM0015060 10100000 319 35.73 6.63 无 No 叶绿体 Chloroplast OePRX36 EVM0017946 0 329 37.29 8.48 有 Yes 细胞质 Cytosol OePRX37 EVM0021366 − 331 36.79 9.34 有 Yes 叶绿体 Chloroplast OePRX38 EVM0060527 0 342 37.64 6.21 有 Yes 细胞质 Cytosol OePRX39 EVM0014494 0 321 35.37 6.18 有 Yes 胞外 Extracellular OePRX40 EVM0042196 0 319 34.12 9.41 有 Yes 叶绿体 Chloroplast OePRX41 EVM0015681 0 346 38.94 8.86 有 Yes 内质网 Endoplasmic reticulum OePRX42 EVM0047610 0 327 35.27 8.80 有 Yes 叶绿体 Chloroplast OePRX43 EVM0047452 20002 379 41.95 8.87 无 No 叶绿体 Chloroplast OePRX44 EVM0061929 0 319 34.23 4.45 有 Yes 胞外 Extracellular OePRX45 EVM0054999 0 291 32.27 5.44 无 No 胞外 Extracellular OePRX46 EVM0048606 0 328 35.67 6.51 有 Yes 液泡膜 Vacular membrane OePRX47 EVM0037058 20 315 34.41 7.55 无 No 内质网 Endoplasmic reticulum OePRX48 EVM0022847 20 413 45.85 9.56 无 No 叶绿体 Chloroplast OePRX49 EVM0020385 20 373 41.16 9.35 无 No 叶绿体 Chloroplast OePRX50 EVM0045080 0 317 34.55 9.02 有 Yes 叶绿体 Chloroplast OePRX51 EVM0044622 0 320 35.53 9.54 有 Yes 叶绿体 Chloroplast OePRX52 EVM0021425 0 324 36.14 6.17 有 Yes 胞外 Extracellular OePRX53 EVM0045494 0 327 35.52 8.71 有 Yes 液泡膜 Vacular membrane OePRX54 EVM0020915 20 306 33.46 8.90 有 Yes 液泡膜 Vacular membrane OePRX55 EVM0031491 0 314 34.07 9.32 有 Yes 叶绿体 Chloroplast OePRX56 EVM0005944 2 292 31.79 9.45 有 Yes 叶绿体 Chloroplast OePRX57 EVM0015088 0 317 34.42 9.13 有 Yes 胞外 Extracellular OePRX58 EVM0050632 0 314 34.04 9.25 有 Yes 叶绿体 Chloroplast OePRX59 EVM0028088 0 317 34.23 8.99 有 Yes 胞外 Extracellular OePRX60 EVM0025336 0 326 34.42 7.58 无 No 叶绿体 Chloroplast OePRX61 EVM0012469 0 317 34.68 9.33 有 Yes 胞外 Extracellular OePRX62 EVM0049574 0 340 37.08 6.25 有 Yes 胞外 Extracellular OePRX63 EVM0050273 10 240 26.55 6.22 无 No 叶绿体 Chloroplast OePRX64 EVM0050817 0 310 34.15 8.46 有 Yes 叶绿体 Chloroplast OePRX65 EVM0009492 0 322 34.85 4.99 有 Yes 叶绿体 Chloroplast OePRX66 EVM0047271 1200201 333 36.96 8.64 无 No 质膜 Plasma membrane OePRX67 EVM0005889 210122 509 58.00 8.73 无 No 细胞核 Nucleus OePRX68 EVM0026529 0 331 36.43 9.50 有 Yes 液泡膜 Vacular membrane OePRX69 EVM0038965 20 310 33.90 9.23 有 Yes 质膜 Plasma membrane OePRX70 EVM0024841 0 328 35.72 8.37 有 Yes 叶绿体 Chloroplast OePRX71 EVM0047959 3 314 34.07 6.94 有 Yes 叶绿体 Chloroplast OePRX72 EVM0059042 0 339 37.43 5.56 有 Yes 质膜 Plasma membrane OePRX73 EVM0022158 0 318 34.18 6.88 有 Yes 叶绿体 Chloroplast OePRX74 EVM0027175 0 326 36.14 9.75 有 Yes 叶绿体 Chloroplast OePRX75 EVM0057531 20 287 30.73 8.89 有 Yes 液泡膜 Vacular membrane OePRX76 EVM0032201 0 293 32.34 8.75 无 No 叶绿体 Chloroplast OePRX77 EVM0059919 0 339 37.61 8.36 有 Yes 液泡膜 Vacular membrane OePRX78 EVM0011789 0 355 39.51 8.06 无 No 质膜 Plasma membrane OePRX79 EVM0024970 − 329 36.54 9.19 有 Yes 叶绿体 Chloroplast OePRX80 EVM0047604 100 339 37.82 8.30 无 No 细胞质 Cytosol OePRX81 EVM0061300 0 330 36.94 5.94 有 Yes 叶绿体 Chloroplast OePRX82 EVM0042525 0 298 32.64 8.62 有 Yes 胞外 Extracellular OePRX83 EVM0045066 0 318 34.03 9.15 有 Yes 叶绿体 Chloroplast OePRX84 EVM0029808 0 354 39.38 5.68 无 No 胞外 Extracellular OePRX85 EVM0032932 1022010 247 27.30 5.65 无 No 细胞质 Cytosol OePRX86 EVM0038819 2001020 250 27.72 5.44 无 No 细胞质 Cytosol OePRX87 EVM0002745 200 321 34.58 4.77 无 No 细胞核 Nucleus OePRX88 EVM0038873 200 326 34.71 8.30 有 Yes 液泡膜 Vacular membrane OePRX89 EVM0026260 2 348 38.22 9.56 有 Yes 质膜 Plasma membrane OePRX90 EVM0044542 0 296 32.61 8.14 有 Yes 胞外 Extracellular OePRX91 EVM0046395 0 318 34.24 9.54 有 Yes 叶绿体 Chloroplast OePRX92 EVM0042595 0 316 33.95 7.51 有 Yes 叶绿体 Chloroplast OePRX93 EVM0047351 0 320 35.08 5.48 有 Yes 叶绿体 Chloroplast OePRX94 EVM0045920 0 332 35.75 4.63 有 Yes 胞外 Extracellular OePRX95 EVM0044144 0 325 35.63 8.75 无 No 叶绿体 Chloroplast OePRX96 EVM0048820 0 314 35.88 8.61 有 Yes 细胞质 Cytosol OePRX97 EVM0058650 0 350 38.67 6.94 有 Yes 叶绿体 Chloroplast OePRX98 EVM0026875 0 371 40.68 6.31 无 No 叶绿体 Chloroplast OePRX99 EVM0019820 0 340 36.88 6.59 有 Yes 胞外 Extracellular OePRX100 EVM0018143 20020200000 394 43.59 9.28 无 No 叶绿体 Chloroplast OePRX101 EVM0029423 0 343 37.01 8.94 有 Yes 叶绿体 Chloroplast OePRX102 EVM0036633 0 342 37.59 9.17 有 Yes 胞外 Extracellular OePRX103 EVM0041887 1 312 33.33 5.97 有 Yes 叶绿体 Chloroplast OePRX104 EVM0032297 0 327 35.71 8.57 有 Yes 胞外 Extracellular OePRX105 EVM0042746 0 302 33.53 8.96 有 Yes 胞外 Extracellular OePRX106 EVM0051394 0 343 37.01 8.94 有 Yes 叶绿体 Chloroplast 2.2 Ⅲ类PRX基因的进化扩张
Ⅲ类PRX基因是一类植物特有的PRX基因,莱茵衣藻(Chlamydomonas reinhardtii)、小立碗藓(Physcomitrella patens)、江南卷柏(Selaginella moellendorffii)[31]、拟南芥[13]、毛果杨[16]、玉米[15]和水稻[14]中分别有6、31、62、73、93、119和138个Ⅲ类PRX基因,说明Ⅲ类PRX基因家族在进化过程中不断发生扩张,这可能与Ⅲ类PRX基因参与了植物广泛的生理过程有关。如图1,利用邻接法构建了包含73个AtPRX、93个PtPRX和106个OePRX蛋白的系统进化树,然后用同样的方法构建了仅包含106个OePRX蛋白的系统进化树,根据3种植物PRX蛋白的系统进化关系和106个OePRX蛋白之间的系统进化关系,将Ⅲ类PRX基因家族分为14个组。组1、2、4聚为1支,组6、7聚为1支,组9单独聚为1支,OePRX和PtPRX基因在这3支中均发生了明显的扩张;组3、10聚为1支,组12单独1支,OePRX和PtPRX基因在这2支中均发生了轻微的收缩。另外,OePRX基因在组13中发生了扩张,而PtPRX基因发生了收缩;PtPRX基因在组5、8、11中发生了扩张,OePRX基因在组5、11中发生了收缩。另外发现:组1 ~ 13中,OePRX、PtPRX、AtPRX基因均有分布;组14中仅包含10个OePRX基因,是一个油橄榄特有的组;这说明组1 ~ 13中,油橄榄、毛果杨、拟南芥有共同的Ⅲ类PRX基因的祖先基因,组14中的OePRX基因发生了明显的遗传分化。
![]() 图 1 油橄榄、毛果杨和拟南芥Ⅲ类PRX蛋白的系统进化关系Figure 1. Phylogeny of class Ⅲ PRX proteins from olive, black cottonwood and Arabidopsis thaliana
图 1 油橄榄、毛果杨和拟南芥Ⅲ类PRX蛋白的系统进化关系Figure 1. Phylogeny of class Ⅲ PRX proteins from olive, black cottonwood and Arabidopsis thaliana2.3 OePRX基因的染色体定位和复制基因分析
2.3.1 OePRX基因在染色体上的不均匀分布
为了分析OePRX基因家族在进化过程中的扩张,绘制了106个OePRX基因在23条染色体上的分布图(图2)。结果发现:100个OePRX基因不均匀地定位于23条染色体上,6个OePRX基因定位于6个不同的contig上。第13条染色体(Chr13)上OePRX基因最多,数量为10(9.4%);其次是第6条染色体,数量为8(7.5%);第10条上有7个OePRX基因(6.6%),第11、12、15、23条上各有6个OePRX基因;第14、19条上各有5个OePRX基因;第3、7、9、17、20、22条上各有4个OePRX基因;第8条上有3个OePRX基因;第1、2、4、5、16、18、21条上各有2个OePRX基因。
![]() 图 2 Ⅲ类油橄榄PRX基因在染色体上的分布图串联复制基因簇用方框标记。Tandem replication gene cluster is marked with a box.Figure 2. Chromosomal distribution of PRX genes in class Ⅲ olive
图 2 Ⅲ类油橄榄PRX基因在染色体上的分布图串联复制基因簇用方框标记。Tandem replication gene cluster is marked with a box.Figure 2. Chromosomal distribution of PRX genes in class Ⅲ olive2.3.2 OePRX共线性分析
油橄榄经历了2次木犀科(Oleaceae)特有的全基因组复制事件,分别发生在约28和59百万年前[1]。为了挖掘OePRX基因家族的进化扩张与油橄榄全基因组复制事件的关系,利用TBtools v1.095中的One Step MCScanX分析了OePRX基因家族内的复制基因,发现OePRX基因家族中共62个OePRX基因参与了基因复制,占OePRX基因总数量的58.5%。如图2所示:15个基因(14.2%)参与了串联复制,共得到了6个串联复制基因簇,6个基因簇分布于不同的染色体上,分别是第6、11、12、13、15、23条染色体。如图3所示:50个基因(47.1%)参与了片段复制,共得到了34对片段复制基因,34对基因分布于19条染色体上,第2、21、22条上未分布,说明片段复制是油橄榄Ⅲ类PRX进化扩张的主要驱动力。值得注意的是,3个基因(OePRX43、OePRX58、OePRX68)既参与了片段复制,也参与了串联复制。
2.3.3 拟南芥、毛果杨和油橄榄PRX基因同线性分析及Ka、Ks分析
为了进一步探究OePRX基因家族的进化扩张,绘制了OePRX与AtPRX和PtPRX基因的同线性图(图4)。结果表明:油橄榄和拟南芥之间有40对Ⅲ类PRX复制基因;油橄榄和毛果杨之间有76对Ⅲ类PRX复制基因。并注意到,25个OePRX(23.6%)基因在拟南芥和毛果杨中均有同源基因,推测在拟南芥、油橄榄和毛果杨分化前,双子叶植物可能有过一次基因复制事件,这与Tuskan等[32]的研究一致。另外,31个OePRX(29.2%)基因在拟南芥中有直系同源基因,46个OePRX(43.4%)基因在毛果杨中有直系同源基因,推测拟南芥与油橄榄的分化可能早于毛果杨与油橄榄的分化。
![]() 图 4 油橄榄、毛果杨和拟南芥Ⅲ类PRX的共线性分析Figure 4. Collinearity analysis of class Ⅲ PRX in olive, black cottonwood and Arabidopsis thaliana
图 4 油橄榄、毛果杨和拟南芥Ⅲ类PRX的共线性分析Figure 4. Collinearity analysis of class Ⅲ PRX in olive, black cottonwood and Arabidopsis thaliana2.3.4 复制基因的Ka、Ks和Ka/Ks值分析
用KaKs_Calculator2.0计算复制基因的Ka、Ks和Ka/Ks值(图5)。在OePRX基因家族的6个串联复制基因簇和34对片段复制基因中,1个串联复制基因簇(OePRX97、OePRX98和OePRX99)和5对片段复制基因(OePRX8/OePRX66、OePRX16/OePRX26、OePRX24/OePRX61、OePRX36/OePRX96、OePRX35/OePRX100)的Ks在0.2 ~ 0.3之间,推测这些基因的复制可能发生于28百万年前的油橄榄全基因复制事件[1]。另外,发现油橄榄复制基因对、Oe-At复制基因对、Oe-Pt复制基因对的Ka/Ks均小于1,说明Ⅲ类PRX在双子叶植物的进化过程中,受到纯化选择的作用。油橄榄片段复制基因对的Ka/Ks小于串联复制基因对的Ka/Ks,说明纯化选择在片段复制基因中更强。
![]() 图 5 油橄榄Ⅲ类PRX各复制基因对的Ka、Ks分析Ka. 非同义替换;Ks. 同义替换;Oe-At. 油橄榄和拟南芥之间的PRX复制基因对;Oe-Pt. 油橄榄和毛果杨之间的PRX复制基因对。Ka, nonsynonymous substitution rate; Ks, synonymous substitution rate; Oe-At, PRX duplicate gene pairs between olive and Arabidopsis thaliana; Oe-Pt, PRX duplicate gene pairs between olive and black cottonwood.Figure 5. Ka, Ks analyses of olive class Ⅲ PRX duplicated genes
图 5 油橄榄Ⅲ类PRX各复制基因对的Ka、Ks分析Ka. 非同义替换;Ks. 同义替换;Oe-At. 油橄榄和拟南芥之间的PRX复制基因对;Oe-Pt. 油橄榄和毛果杨之间的PRX复制基因对。Ka, nonsynonymous substitution rate; Ks, synonymous substitution rate; Oe-At, PRX duplicate gene pairs between olive and Arabidopsis thaliana; Oe-Pt, PRX duplicate gene pairs between olive and black cottonwood.Figure 5. Ka, Ks analyses of olive class Ⅲ PRX duplicated genes2.4 OePRX基因序列分析
2.4.1 PRX基因基序分布和基因结构分析
如图6A,用邻接法构建了106个OePRX蛋白的系统进化树,OePRX蛋白家族分为14个组,组1、2、5、7、14中的OePRX蛋白明显多于其他组。106个OePRX蛋白平均约有324个氨基酸,使用MEME分析了每一个OePRX蛋白的基序分布(图6B),基序个数设置为10,每个基序为6 ~ 50个氨基酸。发现46个OePRX蛋白(43.4%)含有10个基序,34个OePRX蛋白(32.1%)含有9个基序,剩下的OePRX蛋白含有3 ~ 8个基序,说明OePRX蛋白的基序分布整体比较保守。另外还注意到,组1 ~ 13中OePRX蛋白含有6 ~ 10个基序PRX,组14的OePRX蛋白含有3 ~ 5个基序,进一步说明组14中的OePRX蛋白可能发生了功能分化。
![]() 图 6 油橄榄Ⅲ类PRX基因的系统进化树(A)、保守基序(B)和基因结构图(C)Figure 6. Phylogeny (A), conserved motif (B) and extron-intron structure (C) of the olive class Ⅲ PRX genes
图 6 油橄榄Ⅲ类PRX基因的系统进化树(A)、保守基序(B)和基因结构图(C)Figure 6. Phylogeny (A), conserved motif (B) and extron-intron structure (C) of the olive class Ⅲ PRX genes然后,用GSDS绘制了OePRX基因的基因结构图(图6C),OePRX基因中有0 ~ 12个内含子不等,其中,84个OePRX基因(79.2%)中有2 ~ 4个内含子。PRXOePRX基因的大多数内含子位置保守,为0,且所有OePRX基因第1个内含子的位置最为保守,均为0。此外,同组中OePRX基因内含子的数量和位置相对保守,需特别注意的是,组14中OePRX基因中的内含子数量(6 ~ 12个不等)明显多于其他组(0 ~ 4个不等,除OePRX32、OePRX43、OePRX67外),且内含子位置多变,暗示组14中Ⅲ类PRX基因可能具有特殊的进化模式和不同于其他组OePRX基因的功能。
2.4.2 OePRX启动子上顺式作用元件分析
利用OePRX基因转录起始位点上游1 500 bp作为启动子预测顺式作用元件,共预测到59个顺式作用元件(图7)。CAAT-box控制转录起始的频率,TATA-box是RNA聚合酶的结合位点之一,这2个顺式作用元件在106个OePRX基因的启动子中均有分布。34个顺式作用元件(57.6%)与发育有关,其中,光应答顺式作用元件Box 4、G-box、GT1-motif和TCT-motif广泛分布于OePRX基因的启动子中,这说明OePRX基因在油橄榄生长发育过程中具有重要作用。17个顺式作用元件(28.8%)与激素和胁迫应答有关,包括脱落酸(ABA)、水杨酸、赤霉素、茉莉酸甲酯(MeJA)、生长素、低温、干旱、缺氧等,ABRE是依赖ABA表达基因的顺式调节元件,分布于81个OePRX基因的启动子中;CGTCA-motif和TGACG-motif是依赖MeJA表达基因的顺式作用元件,在64个OePRX基因的启动子中有分布;ARE、GC-motif均为厌氧胁迫应答的顺式作用元件,其中ARE分布于76个OePRX基因的启动子中,说明OePRX基因广泛参与激素代谢过程,调控油橄榄的生长发育,且能响应逆境胁迫,增强油橄榄抗逆性。
![]() 图 7 油橄榄Ⅲ类PRX的启动子上的顺式作用元件分析Figure 7. cis-element analysis of promoters of olive class Ⅲ PRX genes
图 7 油橄榄Ⅲ类PRX的启动子上的顺式作用元件分析Figure 7. cis-element analysis of promoters of olive class Ⅲ PRX genes2.5 OePRX表达模式分析
2.5.1 不同器官或组织中OePRX的差异表达
为了研究OePRX基因家族的表达模式,分析了106个OePRX在根、茎、叶、花、果和分生组织中的转录组数据,发现在106个OePRX基因中,61个OePRX基因有表达,tpm在1 ~ 846之间;其余OePRX基因的tpm小于1,可认为其不表达。如图8所示:根据表达模式的相似性,106个OePRX基因在不同组织中的表达量聚为8个模块(module)。18个基因(17.0%,模块A、B和C)在不同组织内普遍表达,其中7个基因(模块A)的整体表达量较高,5个基因(模块B)表达量次之,6个基因(模块C)表达量较低。13个基因(12.3%,模块D、E、F和G)在1个或多个特定的组织内表达,其中,5个基因(模块E和F)在除果实之外的其他组织内表达,2个基因(模块E)在茎、叶、分生组织中表达量更高,7个基因(模块D)主要在花中表达,1个基因OePRX72(模块G)主要在根中表达。值得注意的是,75个基因(70.8%,模块H)几乎不表达或者仅在特定的组织中具有极低的表达。
![]() 图 8 油橄榄Ⅲ类PRX基因在不同器官或组织中的表达模式模块A. 在不同组织中均高表达;模块B. 在不同组织中均有较高表达;模块C. 在不同组织中均较低表达;模块D. 在花中特异高表达;模块E. 在除果外的组织中高表达;模块F. 在除果外的组织较高表达. 模块G. 在根中特异高表达;模块H. 在各组织中低表达或几乎不表达。Module A, high expression in all tissues; module B, relative high expression in all tissues; module C, relative low expression in all tissues; module D, high specific expression in flower; module E, high expression in all tissues except fruit; module F, relative high expression in all tissues except fruit; module G, high specific expression in root; module H, low or no expression in all tissues.Figure 8. Expression patterns of olive class Ⅲ PRX genes in different organs or tissues
图 8 油橄榄Ⅲ类PRX基因在不同器官或组织中的表达模式模块A. 在不同组织中均高表达;模块B. 在不同组织中均有较高表达;模块C. 在不同组织中均较低表达;模块D. 在花中特异高表达;模块E. 在除果外的组织中高表达;模块F. 在除果外的组织较高表达. 模块G. 在根中特异高表达;模块H. 在各组织中低表达或几乎不表达。Module A, high expression in all tissues; module B, relative high expression in all tissues; module C, relative low expression in all tissues; module D, high specific expression in flower; module E, high expression in all tissues except fruit; module F, relative high expression in all tissues except fruit; module G, high specific expression in root; module H, low or no expression in all tissues.Figure 8. Expression patterns of olive class Ⅲ PRX genes in different organs or tissues对比图8中OePRX基因的进化分组和表达模块发现:同一组中的基因表达模式相似,比如组3、7、8、9、10和12中的基因均有单一的基因表达模块;也有一些组中的基因表达模式的变化幅度比较小,比如组2、4、6和11中的基因均聚类为2个表达模块。然而,组5中的基因聚类为6个不同的表达模块,组1、13、14中的基因均聚类为3个不同的表达模块。尽管如此,在组织间普遍表达类模块A、B和C的基因其进化关系更为相近,在特定组织表达类模块D、E、F和G的基因也是如此。另外,值得注意的是,模块H中几乎不表达或者仅在特定的组织表达的75个基因均匀分布于各个组。
2.5.2 实时荧光定量PCR验证
运用实时荧光定量PCR验证了6个基因(OePRX1、OePRX8、OePRX33、OePRX36、OePRX66和OePRX99)在叶片、茎、根和果实中的表达量(图9)。运用Primer Premier5.0设计引物,如表2所示。RNA-seq数据和实时荧光定量结果均表明,OePRX1和OePRX36基因在根中表达量最高,在果实中表达量最低;OePRX8和OePRX66基因在叶片和果实中表达量较高,在茎和根中表达量较低;OePRX33基因在茎和根中表达量较高,在叶和果中表达量较低;OePRX99基因在茎中表达量最高,在果实中表达量最低。
![]() 图 9 RT-qPCR验证转录组数据柱状图为运用RNA-seq得到的tpm值(相对表达量1)。折线图为运用RT-qPCR得到的2−ΔΔCT值(相对表达量2)。Bars indicate tpm value measured by RNA-seq (relative expression 1). Lines indicate 2−ΔΔCT measured by RT-qPCR (relative expression 2).Figure 9. RT-qPCR validations of RNA-seq data表 2 6个油橄榄Ⅲ类PRX基因RT-qPCR的引物Table 2. Primers of six olive class Ⅲ PRX genes used for RT-qPCR
图 9 RT-qPCR验证转录组数据柱状图为运用RNA-seq得到的tpm值(相对表达量1)。折线图为运用RT-qPCR得到的2−ΔΔCT值(相对表达量2)。Bars indicate tpm value measured by RNA-seq (relative expression 1). Lines indicate 2−ΔΔCT measured by RT-qPCR (relative expression 2).Figure 9. RT-qPCR validations of RNA-seq data表 2 6个油橄榄Ⅲ类PRX基因RT-qPCR的引物Table 2. Primers of six olive class Ⅲ PRX genes used for RT-qPCR基因名称
Gene name引物(5′—3′) Primer sequence (5′−3′) 基因产物长度
Gene product length/bpOePRX1 F: TCCAGGAGTTGTTTCTTGTGC 114 R: CCTTCTTCCGTCTTTTCTTCC OePRX8 F: AAGGGCGCATCCTGAAAG 200 R: AGAAGGCATCTTCATCCTTAGC OePRX33 F: GGAAGCATAATAAGCGAGAAG 235 R: TTATTGGAGCCACTCAAACTG OePRX36 F: ACAAACGCCACAAGAACACTG 195 R: CACTCCCTCTCTAAAGCCTCC OePRX66 F: GATTCGCAATGAGGAGGAGTA 140 R: AGCAACAACACCAGCAAGC OePRX99 F: GCTCGGGCTTTTAGAATCATC 204 R: GAAGGAAGGTTTGCCAGTGTT 2.6 非生物胁迫下OePRX基因的差异表达
为了研究油橄榄Ⅲ类PRX基因对非生物胁迫的响应,利用RNA-seq数据分析了油橄榄响应干热胁迫和水涝胁迫过程中差异表达的OePRX基因。如图10所示:干热胁迫下,39个OePRX差异表达,占比36.8%;水涝胁迫下,10个OePRX差异表达,占比9.4%。基于差异表达基因在干热胁迫和复水过程中的表达趋势,将其分为4个模块:模块1,23个基因(21.7%)的表达在干旱胁迫过程中显著下调,复水后显著上调;模块2,7个基因(6.6%)的表达在干旱胁迫过程中显著上调,复水后显著下调;模块3,3个基因(2.8%)的表达在高温和前期干旱时变化不显著,在后期干旱时显著上调,复水后显著下调;模块4,6个基因(5.7%)的表达在高温胁迫下显著上调,解除高温胁迫后,显著下调。另外,与正常浇水的油橄榄相比,在第13天,高温和早期干旱处理的油橄榄中有8个基因(7.5%)显著上调,分别是:OePRX31、OePRX36、OePRX45、OePRX46、OePRX63、OePRX82、OePRX83和OePRX96。在第27天,13个基因(12.3%)显著上调,分别是:OePRX2、OePRX16、OePRX26、OePRX31、OePRX33、OePRX36、OePRX63、OePRX76、OePRX79、OePRX82、OePRX96和OePRX104;4个基因(3.8%)显著下调,分别是OePRX40、OePRX50、OePRX65和OePRX99;在第80天,OePRX基因表达无显著变化。水涝胁迫处理90 d后,油橄榄叶片中10个OePRX基因(9.4%)的表达显著下调,猜测这些差异表达的Ⅲ类PRX在油橄榄响应非生物胁迫胁中具有十分重要的作用。PRX除干热和水淹胁迫外,我国油橄榄常常面临低温和营养等胁迫,因此有必要对其进行更为丰富而广泛地探索,进一步研究Ⅲ类PRX基因在油橄榄响应非生物胁迫过程中的功能。
![]() 图 10 油橄榄Ⅲ类PRX基因响应干热和水涝胁迫的差异表达模式Day(−3). 干旱处理前3天;Day13. 干旱处理第13天且伴随高温;Day27. 干旱处理第27天;Day80. 复水后52天。模块1. 干旱期间基因表达下调,复水后上调;模块2. 干旱期间基因表达上调,复水后下调;模块3. 后期干旱期间基因表达上调,后期干旱及复水后下调;模块4. 仅在高温下基因表达上调,高温胁迫后,基因表达下调。复制基因名称的颜色与系统进化分组的颜色相对应。Day(−3), 3 d before drought treatment; Day13, 13th day of drought treatment with high temperature; Day27, 27th day after drought treatment; Day80, 52 d after re-irrigation. Module 1, gene expression is down regulated during drought and up regulated after rehydration; module 2, gene expression is upregulated during drought and downregulated after rehydration; module 3, gene expression is upregulated during late drought, and downregulated after late drought and rehydration; module 4, gene expression is upregulated only under high temperature, and downregulated after high temperature stress. The color of the replicated gene name corresponds to the color of the phylogenetic grouping.Figure 10. Differential expression patterns of class Ⅲ PRX genes in olive oil in response to dry heat and waterlogging stresses
图 10 油橄榄Ⅲ类PRX基因响应干热和水涝胁迫的差异表达模式Day(−3). 干旱处理前3天;Day13. 干旱处理第13天且伴随高温;Day27. 干旱处理第27天;Day80. 复水后52天。模块1. 干旱期间基因表达下调,复水后上调;模块2. 干旱期间基因表达上调,复水后下调;模块3. 后期干旱期间基因表达上调,后期干旱及复水后下调;模块4. 仅在高温下基因表达上调,高温胁迫后,基因表达下调。复制基因名称的颜色与系统进化分组的颜色相对应。Day(−3), 3 d before drought treatment; Day13, 13th day of drought treatment with high temperature; Day27, 27th day after drought treatment; Day80, 52 d after re-irrigation. Module 1, gene expression is down regulated during drought and up regulated after rehydration; module 2, gene expression is upregulated during drought and downregulated after rehydration; module 3, gene expression is upregulated during late drought, and downregulated after late drought and rehydration; module 4, gene expression is upregulated only under high temperature, and downregulated after high temperature stress. The color of the replicated gene name corresponds to the color of the phylogenetic grouping.Figure 10. Differential expression patterns of class Ⅲ PRX genes in olive oil in response to dry heat and waterlogging stresses3. 讨论与结论
Ⅲ类PRX基因通过氧化特定小分子、调节活性氧水平来促进木质化、细胞伸长、响应生物胁迫和非生物胁迫[33-34]。为了调控基因表达的时空特异性、蛋白分布的特定性和过氧化物酶活性的多样性,高等植物进化得到了庞大的Ⅲ类PRX基因家族[35]。在高等植物的进化过程中,Ⅲ类PRX基因家族通过全基因组复制事件、片段复制、串联复制和转座子复制等方式不断扩张。前人研究表明:拟南芥中有73个PRX基因,9对片段复制基因,10个串联复制基因簇[13];毛果杨中有93个PRX基因,由60 ~ 65百万年前的全基因组复制事件产生了14对复制基因,共有37个串联复制基因簇[16];白梨中有94个PRX基因,26对片段复制基因,1个串联复制基因簇[17];水稻中有138个PRX基因,多于20对的片段复制基因和6个串联复制基因簇[14];玉米中有119个PRX基因,16对片段复制基因和12个串联复制基因簇[15]。本研究中,在双子叶木本植物油橄榄中鉴定得到了106个Ⅲ类PRX基因,其中,62个OePRX(58.5%)参与了基因复制,共产生了6个串联复制基因簇和34对片段复制基因。据此猜想,片段复制可能是高等植物PRX基因家族进化扩张的主要方式。
基因复制事件是基因组进化过程中基因功能冗余和分化的主要来源[36]。复制基因的功能具有相关性,且其功能的分化与其表达模式的分化有关。例如:三倍体小麦中的同源基因VRN-A1、VRN-B1和VRN-D1是调节开花时间的重要因子,不同抽穗期的小麦中,这3个同源基因在不同组织中的表达量不同[37]。本研究发现,许多OePRX基因在不同组织中的表达模式与其在拟南芥或其他模式物种中的表达模式相似,说明PRX的基因功能在不同植物之间具有保守性。AtPRX33和AtPRX34基因是一对高度同源(95%)的Ⅲ类PRX基因,但是其启动子和内含子序列却高度分化。半定量RT-PCR结果表明,这2个基因主要在拟南芥幼苗的根部表达,并且光照对其表达具有促进作用。与野生型拟南芥相比,AtPRX33基因突变体的根变短,AtPRX33和AtPRX34基因双突变体的根更短,而AtPRX34基因过表达拟南芥幼苗的根显著变长[38]。本研究发现:OePRX基因家族中的6个串联复制基因簇和34个片段复制基因的分子量、等电点、信号肽的分布和长度以及基序分布均比较保守;但2个串联复制基因簇(33.3%)和22对片段复制基因(64.7%)的内含子位置发生分化;1个串联复制基因簇(16.7%)和11对片段复制基因(32.4%)在不同组织中的表达模式发生分化。因此,有必要利用分子生物学手段深入研究油橄榄Ⅲ类PRX基因家族内复制基因的亚功能和新功能。
为了研究单个Ⅲ类PRX基因在逆境下的特定功能,大量Ⅲ类PRX转基因植株已经获得。过表达甜土豆(Ipomoea batatas)swpa4 PRX基因的烟草(Nicotiana tabacum)幼苗和过表达AtPRX3基因的拟南芥幼苗[39]的耐盐性和耐旱性增强。过表达AtPRX22、AtPRX39和AtPRXrx69基因的拟南芥对油菜素类固醇不敏感,而突变体的耐寒性增强[40]。同样地,2个长春花(Catharanthus roseus)PRX基因在烟草中的异源转化提高了其在脱水胁迫下的发芽率,同时提高了其耐旱性[41]。辣椒(Capsicum annuum)CpPRX2的转基因拟南芥幼苗提高了其体内H2O2水平的积累,同时提高了其耐病性、耐旱性和耐盐性[42]。菜豆(Phaseolus vulgaris)过氧化物酶FBP1反义表达的拟南芥中,AtPRX33和AtPRX34基因的表达量降低,体内氧化应激受到抑制,抗病性减弱[43]。本研究发现:干热胁迫和水涝胁迫下油橄榄中40个OePRX基因(37.7%)显著差异表达,且差异表达模式多样(图10),猜测其他OePRX基因可能参与另外的生物胁迫或非生物胁迫,需要对多种胁迫下的油橄榄幼苗开展更为广泛的研究,锁定响应胁迫的关键基因,从而深入挖掘其功能。Wu等[44]研究表明:经历了多次多倍化复制的植物保留了许多胁迫相关的基因家族,且得到的复制基因通常是胁迫相关通路的关键因子。本研究结果表明:与正常生长的油橄榄相比,干热胁迫下油橄榄中串联复制基因簇(OePRX58和OePRX59)和3对片段复制基因(OePRX16和OePRX26,OePRX36和OePRX96,OePRX37和OePRX79)的表达量均显著降低,片段复制基因对(OePRX22和OePRX64)的表达量均显著上升,片段复制基因对(OePRX1和OePRX45)的表达量仅在高温下显著上升;水涝胁迫下,片段复制基因对(OePRX36和OePRX96)的表达量均显著降低,猜测这些复制基因之间具有较强的功能相关性,可能是研究油橄榄非生物胁迫响应通路的关键候选基因。
综上所述,油橄榄OePRX基因家族明显的进化扩张和多变的表达模式暗示了其功能的复杂性,尤其是复制基因的亚功能化和新功能化对油橄榄在地中海地区的广泛分布具有十分重要的适应性意义。
-
![]()
图 1 毛乌素沙地行政区划及周围气象台站空间分布
Figure 1. Administrative division of the Mu Us Desert and spatial distribution of weather stations around it
![]()
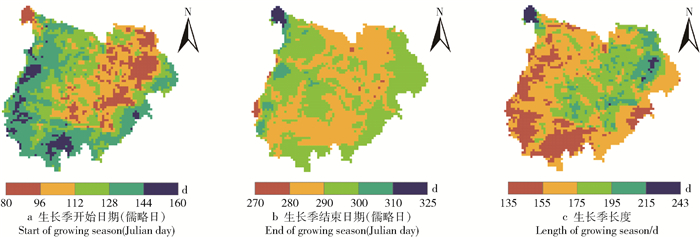
图 3 1982—2015年毛乌素沙地植被平均物候的空间分布
Figure 3. Spatial distribution of mean vegetation phenology in the Mu Us Desert from 1982 to 2015
![]()
图 4 1982—2015年毛乌素沙地物候的年际变化
Figure 4. Inter-annual variations of phenology in the Mu Us Desert from 1982 to 2015
![]()
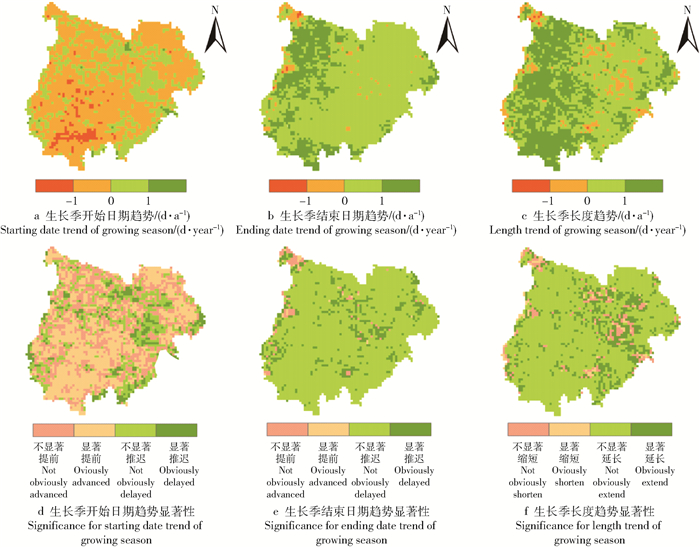
图 5 1982—2015年毛乌素沙地物候期动态变化
Figure 5. Dynamic change in phenology in the Mu Us Desert from 1982 to 2015
![]()
图 6 毛乌素沙地植被物候与气象因子的关系
Figure 6. Relationship between phenology and meteorological factors in the Mu Us Desert
![]()
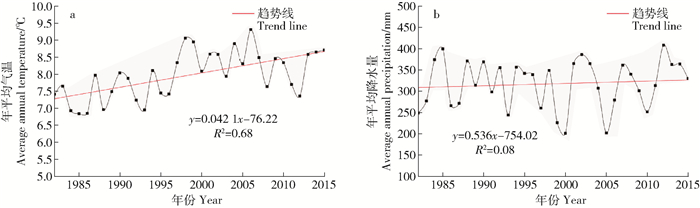
图 7 毛乌素沙地年平均气温和降水量的年际变化
Figure 7. Interannual changes of average annual temperature and precipitation in Mu Us Desert
表 1 本文物候结果与其他研究结果的比较
Table 1 Comparison of phenology results in this study and previous studies
研究区域
Study area生长季开始日期
(儒略日)
Beginning date of growing season
(Julian day)/d生长季结束日期
(儒略日)
Ending date of growing season
(Julian day)/d生长季长度
Length of growing season/d研究时段
Study period数据及分辨率
Data and resolution ratio文献来源
Source of literature毛乌素沙地
Mu Us Desert80~160 270~325 135~243 1982—2015 AVHRR LTDR-V4 (5km) 本文
This study陕甘宁交界*
Shaanxi-Gansu-Ningxia junction80~160 290~330 130~250 1999—2010 SPOT-VGT (1km) 韦振锋等[29]
Wei Z F, etc[29]黄土高原*
The Loess Plateau100~145 283~305 135~205 1982—2011 AVHRR LTDR-V4 (5km) 谢宝妮等[ 17]
Xie B N, etc [ 17]毛乌素沙地
Mu Us Desert90~156 245~323 125~200 2001—2013 MODIS (0.25km) 王静璞等[ 22]
Wang J P, etc [ 22]中国北方*
North of China92~142 290~300 125~200 1982—2006 AVHRR GIMMS (8km) 安佑志等[ 28]
An Y Z, etc [ 58]注:*表示选择文献中的研究区域与本文研究区域重合的区域。Note: * represents selecting the area in the literature that overlaps the study area in the paper.  下载: 导出CSV
下载: 导出CSV
表 2 生长季开始日期向前累计0~5个月的气温和降水量的年际变化
Table 2 Interannual changes in cumulative 0-5 months air temperature and precipitation prior to the beginning date of the growing season
月的个数
Number of month气温的趋势/(℃·a -1)
Trend of air temperature/(℃·year -1)降水量的趋势/(mm·a -1)
Trend of precipitation/(mm·year -1)0 0.044 * -0.13 1 0.041 * 0.41 2 0.036 * 0.49 3 0.038 * 0.56 4 0.033 * 0.79 5 0.043 * 0.34 注:*表示在0.05的水平上通过显著性检验。下同。Notes: * stands for passing significant test at the 0.05 level. The same below.
下载: 导出CSV
表 3 生长季结束日期向前累计0~5个月的气温和降水量的年际变化
Table 3 Interannual changes in cumulative 0-5 months air temperature and precipitation prior to the ending date of the growing season
月的个数
Number of month气温的趋势/(℃·a -1)
Trend of air temperature/(℃·year -1)降水量的趋势/(mm·a -1)
Trend of precipitation/(mm·year -1)0 0.054 * 0.64 1 0.047 * 0.34 2 0.045 * 0.54 3 0.051 * 0.21 4 0.039 * -0.19 5 0.045 * 0.52
下载: 导出CSV
-
[1] Wan M W, Liu X Z.Observation method of the phenology in China[M]. Beijing: Science Press, 1979:1.
[2] Cong N, Piao S, Chen A, et al. Spring vegetation green-up date in China inferred from SPOT NDVI data: a multiple model analysis[J]. Agricultural and Forest Meteorology, 2012, 165:104-113. doi: 10.1016/j.agrformet.2012.06.009
[3] Piao S L, Fang J Y, Zhou L M, et al. Variations in satellite-derived phenology in China's temperate vegetation[J]. Global Change Biology, 2010, 12(4):672-685. http://cn.bing.com/academic/profile?id=59cdfdb4db3cdd16879116ff2c8c0a17&encoded=0&v=paper_preview&mkt=zh-cn
[4] Miao G S, Geng X Z, Nan C, et al. Increasing altitudinal gradient of spring vegetation phenology during the last decade on the Qinghai-Tibetan Plateau[J]. Agricultural and Forest Meteorology, 2014, 189:71-80. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=48d15a73cb0e805043a6ed42f0a61866
[5] 许青云, 杨贵军, 龙慧灵, 等.基于MODIS NDVI多年时序数据的农作物种植识别[J].农业工程学报, 2014, 30(11):134-144. doi: 10.3969/j.issn.1002-6819.2014.11.017 Xu Q Y, Yang G J, Long H L, et al. Crop information identification based on MODIS NDVI time-series data[J]. Transactions of the Chinese Society of Agricultural Engineering, 2014, 30(11): 134-144. doi: 10.3969/j.issn.1002-6819.2014.11.017
[6] 陈静茹, 杜彦君, 张玉红, 等.黑龙江木本植物盛花期对气候变暖的响应[J].北京林业大学学报, 2016, 38(11):50-56. doi: 10.13332/j.1000-1522.20160186 Chen J R, Du Y J, Zhang Y H, et al, Response of woody plant flowering season to climate warming in Heilongjiang[J]. Journal of Beijing Forestry University, 2016, 38(11):50-56. doi: 10.13332/j.1000-1522.20160186
[7] 康峻, 牛铮, 高帅, 等.基于拟合物候参数的植被遥感决策树分类[J].农业工程学报, 2014, 30(9):148-156. doi: 10.3969/j.issn.1002-6819.2014.09.019 Kang J, Niu Z, Gao S, et al. Decision tree classification based on fitted phenology parameters from remotely sensed vegetation data[J]. Transactions of the Chinese Society of Agricultural Engineering, 2014, 30(9): 148-156. doi: 10.3969/j.issn.1002-6819.2014.09.019
[8] 宋富强, 康慕谊, 杨朋, 等.陕北地区GIMMS、SPOT-VGT和MODIS归一化植被指数的差异分析[J].北京林业大学学报, 2010, 32(4):72-80. http://j.bjfu.edu.cn/article/id/9419 Song F Q, Kang M Y, Yang P, et al. Analysis of GIMMS, SPOT-VGT and MODIS normalized difference vegetation index in Northern Shanxi[J]. Journal of Beijing Forestry University, 2010, 32(4):72-80. http://j.bjfu.edu.cn/article/id/9419
[9] 王鸽, 韩琳, 张昱.东北地区地表NDVI的时空变化规律[J].北京林业大学学报, 2012, 34(6):86-91. http://j.bjfu.edu.cn/article/id/9846 Wang G, Han L, Zhang Y. Temporal and spatial variation of surface NDVI in Northeast China[J]. Journal of Beijing Forestry University, 2012, 34(6):86-91. http://j.bjfu.edu.cn/article/id/9846
[10] Guo L, Dai J, Wang M, et al. Responses of spring phenology in temperate zone trees to climate warming: a case study of apricot flowering in China[J]. Agricultural and Forest Meteorology, 2015, 201:1-7. http://cn.bing.com/academic/profile?id=6df719d9343ed8818e3966e81c6fc90b&encoded=0&v=paper_preview&mkt=zh-cn
[11] 刘峻明, 李曼曼, 王鹏新, 等.基于LAI时间序列重构数据的冬小麦物候监测[J].农业工程学报, 2013, 29(19):120-129. doi: 10.3969/j.issn.1002-6819.2013.19.015 Liu J M, Li M M, Wang P X, et al. Monitoring of phenology by reconstructing LAI time series data for winter wheat[J]. Transactions of the Chinese Society of Agricultural Engineering, 2013, 29(19): 120-129. doi: 10.3969/j.issn.1002-6819.2013.19.015
[12] Heumann B W, Seaquist J W, Eklundh L, et al. AVHRR derived phenological change in the Sahel and Soudan, Africa, 1982-2005[J]. Remote Sensing of Environment, 2007, 108(4):385-392. doi: 10.1016/j.rse.2006.11.025
[13] Lee R, Yu F, Price K P, et al. Evaluating vegetation phenological patterns in Inner Mongolia using NDVI time-series analysis[J]. International Journal of Remote Sensing, 2002, 23(12): 2505-2512. doi: 10.1080/01431160110106087
[14] Kim Y, Kimball J S, Didan K, et al. Response of vegetation growth and productivity to spring climate indicators in the conterminous United States derived from satellite remote sensing data fusion[J]. Agricultural and Forest Meteorology, 2014, 194:132-143. doi: 10.1016/j.agrformet.2014.04.001
[15] Menzel A, Sparks T H, Estrella N, et al. European phenological response to climate change matches the warming pattern[J]. Global Change Biology, 2006, 12(10):1969-1976. doi: 10.1111/gcb.2006.12.issue-10
[16] 宋春桥, 游松财, 柯灵红, 等.藏北高原植被物候时空动态变化的遥感监测研究[J].植物生态学报, 2011, 35(8):853-863. doi: 10.1079-SSR2002117/ Song C Q, You S C, Ke L H, et al. Study on temporal and spatial dynamic changes of vegetation phenology in northern Tibet by remote sensing[J]. Chinese Journal of Plant Ecology, 2011, 35(8):853-863. doi: 10.1079-SSR2002117/
[17] 谢宝妮, 秦占飞, 王洋, 等.基于遥感的黄土高原植被物候监测及其对气候变化的响应[J].农业工程学报, 2015, 31(15):153-160. doi: 10.11975/j.issn.1002-6819.2015.15.021 Xie B N, Qin Z F, Wang Y, et al. Monitoring vegetation phenology and their response to climate change on Chinese Loess Plateau based on remote sensing[J]. Transactions of the Chinese Society of Agricultural Engineering, 2015, 31(15): 153-160. doi: 10.11975/j.issn.1002-6819.2015.15.021
[18] 王涛.中国沙漠化研究的进展[J].中国沙漠, 1999, 19(4):299-311. doi: 10.3321/j.issn:1000-694X.1999.04.001 Wang T. Advances in desertification research in China[J]. Journal of Desert Research, 1999, 19 (4): 299-311. doi: 10.3321/j.issn:1000-694X.1999.04.001
[19] 北京大学地理系.毛乌素沙区自然条件及其改良利用[M].北京:科学出版社, 1983. Department of Geography, Peking University. Mu Us sandy natural conditions and its improved utilization[M]. Beijing: Science Press, 1983.
[20] 侯学会, 牛铮, 高帅, 等.基于SPOT-VGTNDVI时间序列的农牧交错带植被物候监测[J].农业工程学报, 2013, 29(1):142-150. http://www.cnki.com.cn/Article/CJFDTotal-NYGU201301021.htm Hou X H, Niu Z, Gao S, et al. Monitoring vegetation phenology in farming-pastoral zone using SPOT-VGT NDVI data[J]. Transactions of the Chinese Society of Agricultural Engineering, 2013, 29(1): 142-150. http://www.cnki.com.cn/Article/CJFDTotal-NYGU201301021.htm
[21] 马勇刚, 张弛, 塔西甫拉提·特依拜.中亚及中国新疆干旱区植被物候时空变化[J].气候变化研究进展, 2014, 10(2):95-102. doi: 10.3969/j.issn.1673-1719.2014.02.003 Ma Y G, Zhang C, Tassilati·Teyibai. Spatial and temporal variation of vegetation phenology in arid zones in Xinjiang, central Asia and China[J]. Climate Change Research, 2014, 10(2): 95-102. doi: 10.3969/j.issn.1673-1719.2014.02.003
[22] 王静璞, 刘连友, 贾凯, 等.毛乌素沙地植被物候时空变化特征及其影响因素[J].中国沙漠, 2015, 35(3):624-631. http://d.old.wanfangdata.com.cn/Periodical/zgsm201503014 Wang J P, Liu L Y, Jia K, et al. Spatiotemporal variation of vegetation phenology and its affecting factors in the Mu Us Sandy Land[J]. Journal of Desert Research, 2015, 35(3):624-631. http://d.old.wanfangdata.com.cn/Periodical/zgsm201503014
[23] 王涛.中国北方沙漠与沙漠化图集[M].北京:科学出版社, 2014. Wang T. Atlas of desertification and desertification in northern China[M]. Beijing: Science Press, 2014.
[24] Pedelty J, Devadiga S, Masuoka E, et al. Generating a long-term land data record from the AVHRR and MODIS instruments[C]//Geoscience and Remote Sensing Symposium, 2007. IEEE International. Barcelona: IEEE, 2008: 1021-1025.
[25] Holben B N. Characteristics of maximum-value composite images from temporal AVHRR data[J]. International Journal of Remote Sensing, 1986, 7(11):1417-1434. doi: 10.1080/01431168608948945
[26] Jakubauskas M E. Harmonic analysis of time-series AVHRR NDVI data[J]. Photogrammetric Engineering and Remote Sensing, 2001, 67(4):461-470. http://cn.bing.com/academic/profile?id=cc5ccd585c177db84a28fbcf2d7a3be1&encoded=0&v=paper_preview&mkt=zh-cn
[27] Sha Z Y, Zhong J L, Bai Y F, et al. Spatio-temporal patterns of satellite-derived grassland vegetation phenology from 1998 to 2012 in Inner Mongolia, China[J]. Journal of Arid Land, 2016, 8(3):462-477. doi: 10.1007/s40333-016-0121-9
[28] 安佑志.基于遥感的中国北部植被NDVI和物候变化研究[D].上海: 华东师范大学, 2014. An Y Z. Remote sensing based NDVI and phenological changes in northern China[D]. Shanghai: East China Normal University, 2014.
[29] 韦振锋, 任志远, 张翀, 等. 1999—2010年陕甘宁黄土高原区气候对植被物候的影响[J].水土保持通报, 2014, 34(5):232-236. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=stbctb201405045 Wei Z F, Ren Z Y, Zhang J, et al. Impact of climate on vegetation phenology in the Loess Plateau region of Shaanxi, Gansu, and Ningxia in 1999-2010[J]. Bulletin of Soil and Water Conservation, 2014, 34(5):232-236. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=stbctb201405045
[30] Ma T, Zhou C H. Climate-associated changes in spring plant phenology in China[J]. International Journal of Biometeorology, 2012, 56(2): 269-275. doi: 10.1007/s00484-011-0428-3
[31] Wei H Y, Heilman P, Qi J G, et al. Assessing phenological change in China from 1982 to 2006 using AVHRR imagery[J]. Frontiers of Earth Science, 2012, 6(3): 227-236. doi: 10.1007/s11707-012-0321-3
[32] Wang H, Liu D, Lin H, et al. NDVI and vegetation phenology dynamics under the influence of sunshine duration on the Tibetan Plateau[J]. International Journal of Climatology, 2015, 35(5):687-698. doi: 10.1002/joc.2015.35.issue-5
[33] Zhang X, Friedl M A, Schaaf C B, et al. Monitoring vegetation phenology using MODIS[J]. Remote Sensing of Environment, 2003, 84(3):471-475. doi: 10.1016/S0034-4257(02)00135-9
[34] Piao S L, Cui M, Chen A, et al. Altitude and temperature dependence of change in the spring vegetation green-up date from 1982 to 2006 in the Qinghai-Xizang Plateau[J]. Agricultural and Forest Meteorology, 2011, 151(12):1599-1608. doi: 10.1016/j.agrformet.2011.06.016
[35] 闫峰, 吴波.近40a毛乌素沙地荒漠化过程研究[J].干旱区地理, 2013, 36(6):987-996. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=ghqdl201306003 Yan F, Wu B. Study on the desertification process in Mu Us Desert in recent 40 years[J]. Arid Land Geography, 2013, 36(6):987-996. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=ghqdl201306003
[36] Schwalm C R, Anderegg W R L, Michalak A M, et al. Global patterns of drought recovery[J]. Nature, 2015, 548:202-205. http://d.old.wanfangdata.com.cn/OAPaper/oai_pubmedcentral.nih.gov_3419014
[37] 谢宝妮.黄土高原近30年植被覆盖变化及其对气候变化的响应[D].杨凌: 西北农林科技大学, 2016. Xie B N. Changes of vegetation coverage and its response to climate change in the Loess Plateau in recent 30 years[D]. Yangling: Northwest A&F University, 2016.
-
期刊类型引用(3)
1. 吴昊,姜俊慧,罗莉,崔益萌,张婉莹. UV-B辐射增强和氮添加及其互作对空心莲子草-马唐竞争的影响. 西南农业学报. 2024(02): 344-352 .  百度学术
百度学术
2. 吴昊,谢闪闪,苏仪函. 氮添加对茶树与入侵杂草空心莲子草种间竞争的影响. 南方农业学报. 2024(05): 1335-1346 . 百度学术
3. 吴昊,刘雨欣,李利军,余梅鹏,杨依琳,庄启国,杜奎. 人为干扰对猕猴桃园杂草多样性与土壤因子关系的影响. 西南农业学报. 2024(06): 1313-1321 . 百度学术
其他类型引用(0)














计量
- 文章访问数: 3222
- HTML全文浏览量: 763
- PDF下载量: 93
- 被引次数: 3
