
Changing patterns of nitrogen and phosphorus contents in leaf-branch-root of natural Pinus tabuliformis with precipitation and temperature
-
摘要:目的
对油松叶、枝、根的氮、磷含量随水热变化的响应规律进行研究,有助于加深对我国暖温带地区森林植物应对气候变化策略的认识。
方法本文基于生态化学计量学理论,在我国天然油松林主要分布区选择7个代表性样点,对优势种油松叶、枝、根的氮、磷含量进行定量分析。
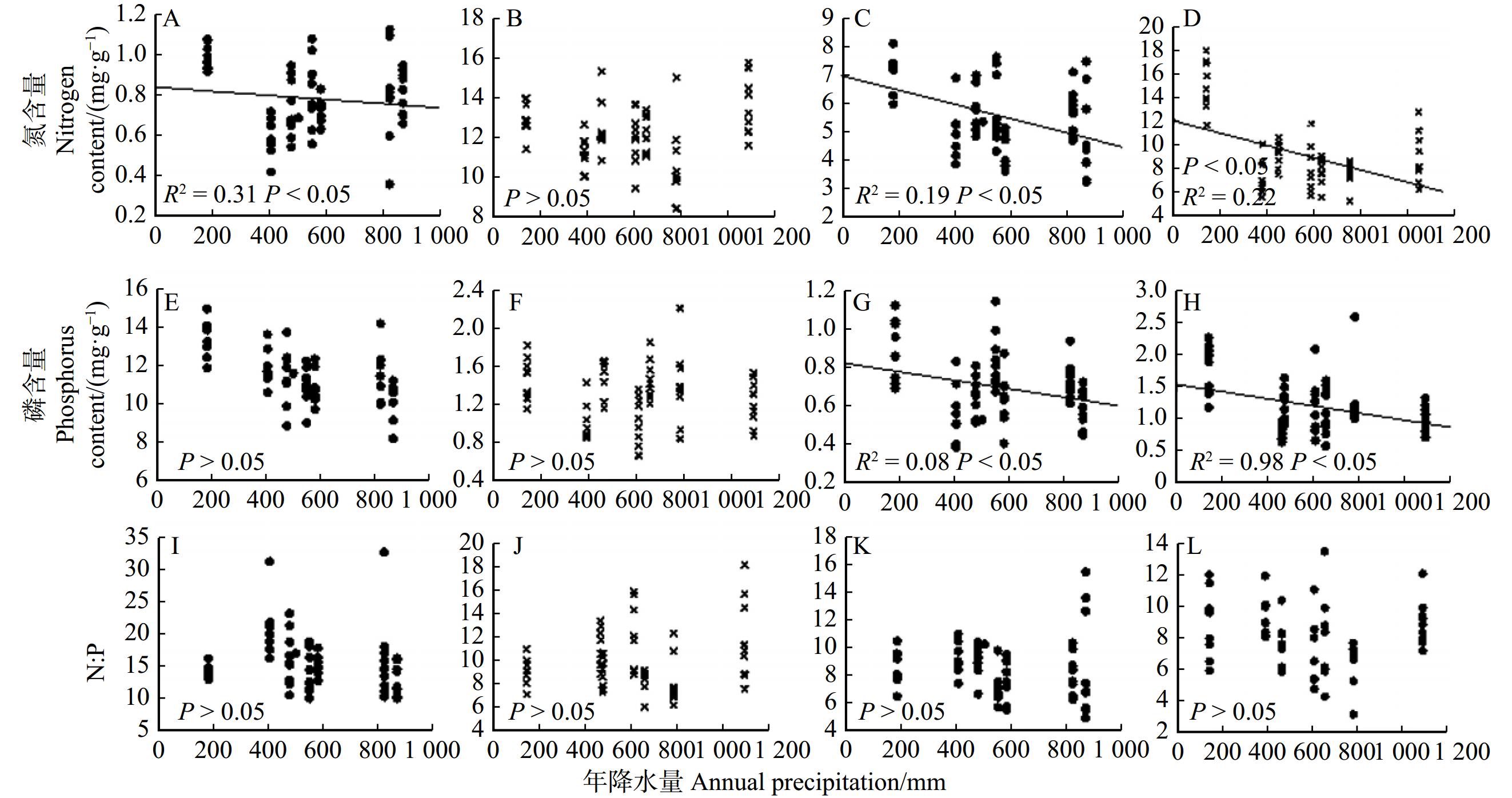
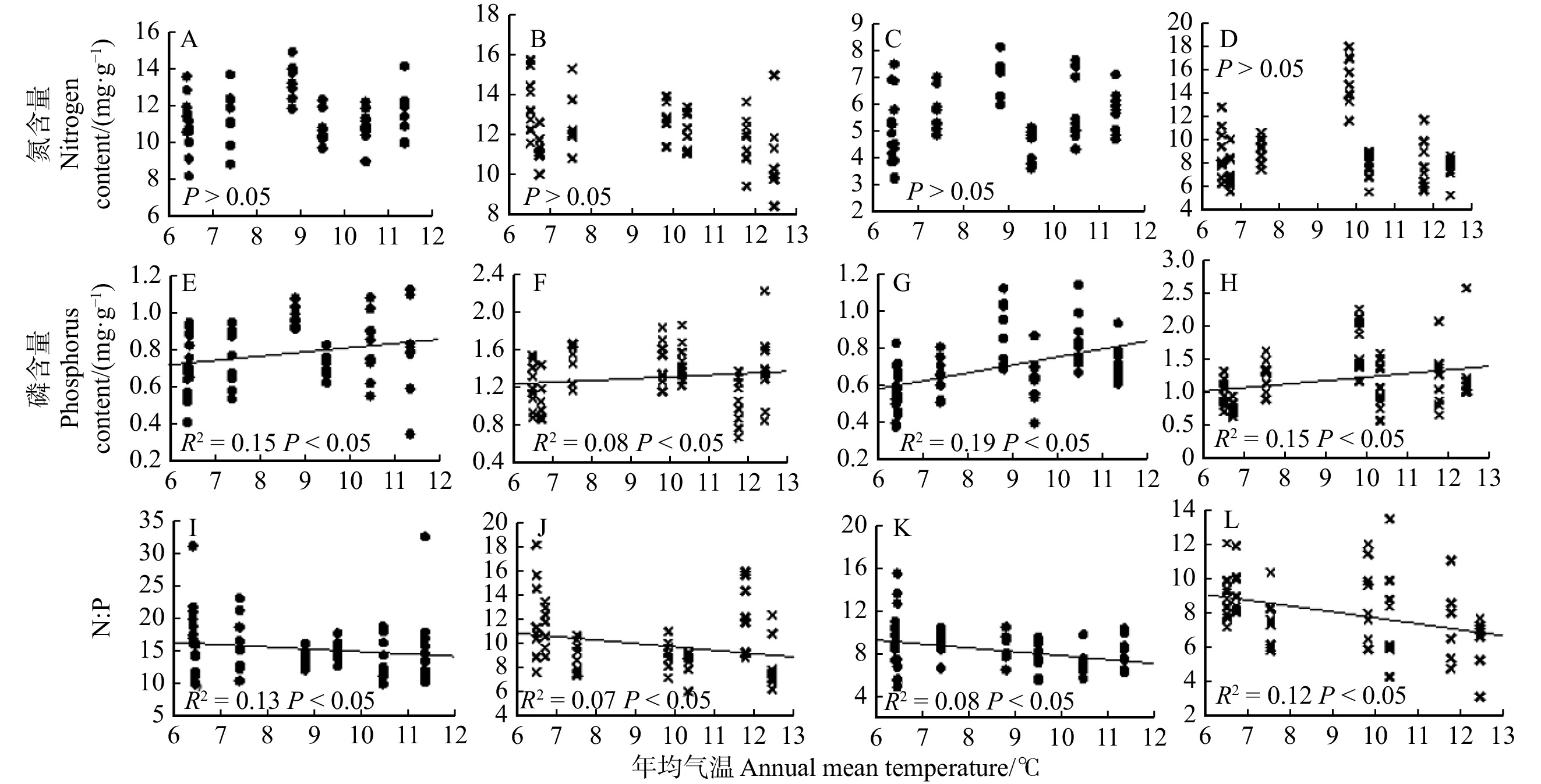
结果(1)油松叶、枝、根器官间氮、磷含量存在显著差异,叶的氮、磷含量均高于枝和根的氮、磷含量,其中当年生叶、当年生枝的氮、磷含量高于多年生叶、多年生枝的氮、磷含量。吸收根氮、磷含量高于次生根氮、磷含量。当年生叶、当年生枝的氮磷比低于多年生叶、多年生枝的氮磷比,次生根氮磷比高于吸收根氮磷比。(2)油松叶、枝、根的氮、磷含量的变异程度存在差异,整体变异系数范围为12.3% ~ 44.4%,其中吸收根磷含量的变异系数最大,多年生叶氮含量的变异系数最小。(3)油松不同器官的氮磷化学计量特征随环境变化的响应存在差异,油松多年生叶氮含量、多年生及当年生枝氮、磷含量随降水量的增加呈显著下降的趋势。多年生及当年生叶磷含量、枝磷含量均随温度的升高呈显著上升的趋势。多年生及当年生叶氮磷比、枝氮磷比随温度的升高呈显著下降的趋势。次生根磷含量随温度的升高呈显著上升的趋势,次生根氮磷比随温度升高呈显著下降的趋势。
结论降水、温度对我国天然油松林优势物种油松叶、枝、根的氮磷化学计量特征有显著影响,不同器官应对这两个环境因子的响应趋势及适应机制不同。
Abstract:ObjectiveStudying the response of nitrogen and phosphorus contents in leaf-root of Pinus tabuliformis to hydrothermal changes can enhance our understanding of the strategies for forest community plants in warm temperate regions of China to confront the challenges posed by climatic shifts.
MethodSeven representative sample locations were selected in the main distribution areas of natural P. tabuliformis forests in China, and the nitrogen (N) and phosphorus (P) contents in the leaves, branches, and roots of P. tabuliformis were meticulously assessed according to the theory of ecological stoichiometry.
Result(1) Differences in nitrogen (N) and phosphorus (P) contents among various plant organs of leaves, branches, and roots were evident. Notably, concentrations of N and P in leaves surpassed those in branches and roots. Additionally, both current leaves and branches exhibited higher N and P levels compared with their perennial counterparts, while absorbed roots displayed elevated N and P contents compared with secondary roots. The N∶P ratios in current leaves and branches were lower than those in perennial leaves and in perennial branches, whereas secondary roots demonstrated a higher N∶P ratio than absorbed roots. (2) Variability in nitrogen (N) and phosphorus (P) content differed among leaves, branches, and roots, as indicated by the overall coefficient of variation ranging from 12.3% to 44.4%. Notably, the highest variability was observed in absorbed roots, contrasting with the lowest variability observed in perennial leaves. (3) The adaptive responses of nitrogen (N) and phosphorus (P) stoichiometry within distinct organs of P. tabuliformis to environmental shifts exhibited notable variations. Specifically, the N content in perennial leaves, perennial branches, and current branches, along with the P content in perennial branches and both current branches and leaves, experienced significant decreases with escalating precipitation. Conversely, a significant uptrend in P content was observed in perennial leaves, perennial branches, current leaves, and current branches as the temperature increased. Simultaneously, the N∶P ratio in perennial leaves, perennial branches, and both current leaves and branches demonstrated substantial declines with rising temperatures. Notably, the P content in secondary roots displayed a marked increase with elevated temperature, whereas the N∶P ratio in secondary roots exhibited a notable decrease.
ConclusionBoth precipitation and temperature exert significant impacts on the nitrogen (N) and phosphorus (P) stoichiometry in the leaves, branches, and roots of P. tabuliformis. The diverse response trends and adaptation mechanisms of distinct organs to these two environmental factors exhibite marked variations.
-
气候变化已经成为21世纪人类共同面临的重大环境问题,大气中CO2等温室气体浓度的增加,致使全球气候系统正在发生着有史以来从未有过的急剧变化,主要表现在全球变暖、降水格局变化、海平面上升等方面[1]。全球变暖的影响在北半球中高纬度的大陆内部表现的尤为明显[2]。森林作为陆地生态系统的重要组成部分,一直以来都是气候变化研究中的重点对象[3]。东北地区是我国纬度最高的区域,该区域内的森林对气候变化的响应受到全球气候变暖的影响,还具有明显的地域性特点[4]。众多研究表明,温度、降水对该地区的森林都有一定的影响[5-7],其中高温造成的负面影响尤为显著[8]。
长白落叶松(Larix olgensis)是北半球高纬度地区分布较为广泛的针叶树种之一,在海拔500~1 950 m均有分布并常形成大面积纯林。国外已有不少针对落叶松相关种属的生长与气候变化关系的研究在与我国临近的西伯利亚地区开展[9-12]。我国已有较多关于东北地区长白落叶松对气候变化响应的研究[13-15],但大多集中于长白山一带且多是针对高海拔或林线处林分[13, 16-17],在其他区域的研究还相对较少。吉林北部山区接近长白落叶松在我国的分布北界[18],在该区域开展研究对理解长白落叶松林对气候变化的响应有着重要的意义[19]。同时,该区域气候干暖化趋势明显,对树木生长已经产生了显著影响[20],急需开展相关研究为该地区森林的经营和保护提供理论依据。
树木生长对气候变化的响应受到树种、气候及林分因子等多方面的影响。如Brooks等[21]提出气候变化对不同树种的影响有差异,温度上升可能严重地影响某一树种的生长,却对另一树种影响不大。针对长白落叶松、兴安落叶松(L. gmelinii)进行的研究发现,不同海拔(温度、降水梯度)的落叶松对气候变化的响应存在明显差异[13, 22]。除树种、气候之外,还存在其他因素的影响。例如不同树龄的兴安落叶松的生长对气候变化的响应存在明显差异,老龄林更易受到水分胁迫的影响[23]。因此,气候条件、林分因子对森林与气候变化之间的响应关系都存在影响,但鲜有研究同时探究二者共同的影响及其各自的相对作用大小。
目前有关森林对气候变化响应的研究,多采用植被覆盖指数(Normalized difference vegetation index, NDVI)、碳收支、生产力等指标作为反映[5-6]。而用树木年轮(简称树轮)来研究生态系统对气候变化的响应,是全球气候变化研究的热点[24-26],相对其他方法,具有定年准确、连续性强、分辨率高等优点[27]。研究树木径向生长与气候因子的关系有助于理解一个地区森林生产力的生态限制因素,以及气候变化条件下森林生态系统的变化机理[8-9]。
本文以吉林北部山区不同林龄的长白落叶松天然林为研究对象,通过研究树木径向生长与历史气候变化的关系,探讨了气候条件、林分因子(林龄、林分密度、蓄积量、立地条件等)对年轮—气候关系的影响,以明确影响长白落叶松林对气候变化响应方式的主要影响因素。这对于在未来全球气候变暖的大背景下为该地区的长白落叶松林提供合理的保护抚育措施具有参考价值。
1. 研究区概况
研究区为吉林省北部山区,位于长白山北部余脉至老爷岭之间(42°49′48″~43°31′12″ N、128°00′36″~130°41′24″ E),沿纬度梯度自南向北包括敦化市的大蒲柴河林场(DPC)、汪清县的金苍林场(JC)和六道林场(LD)等3个研究地点,海拔550~800 m。研究区内为典型的温带季风气候,具有冬季冷且长、夏季热而短且四季分明的特点,年均气温1.0~3.0 ℃,最冷月均温-18 ℃,年降水量690~750 mm。该地区地带性植被为阔叶红松(Pinus koraiensis)林(分布于海拔500~1 100 m),也分布着各种次生森林类型,以及长白落叶松等人工林[28]。研究区内的长白落叶松林混生有少量白桦(Betula platyphylla)、春榆(Ulmus davidiana)、蒙古栎(Quercus mongolica)等阔叶树种,同时还伴生有少量欧亚大陆针叶林中的寒温性树种,如鱼鳞云杉(Picea jezonensis var. microsperma)、红皮云杉(Picea koraiensis)等。土壤以棕黑色的壤土或砂壤土为主。
2. 研究方法
2.1 树木年轮采样与年表建立
野外调查于2016年8—9月份进行,在上述3个研究地点分别设置5、4和8个20 m×50 m的长白落叶松样地(表 1)。对样地中所有胸径大于3 cm的乔木个体挂牌并进行每木检尺,测量树高、冠幅等。各样地的林龄采取国内外相关研究中常用的方法进行估测[29],使用生长锥钻取样地中胸径最大的5株乔木的完整年轮样品(至髓心),以这5个个体的平均年龄作为该样地的林龄,这样可以防止出现由于个别个体树龄较大而导致对林分年龄的估计偏高。另外,蓄积量采用相关蓄积量方程计算得到,立地条件则参考土壤表层含水率并结合实地调查确定,划分为好、中、差3个等级。
表 1 样地基本信息表Table 1. Basic information of each sample plot样地号
Sample plot纬度
Latitude经度
Longitude林龄
Stand age/a立地条件
Site
condition林分密度/(株·hm-2)
Stand density/
(tree·ha-1)蓄积量/(m3·hm-2)
Volume/
(m3·ha-1)平均树高
Mean tree
height/m平均胸径
Mean DBH/
cmDPC1 42°51′N 128°04′E 31 中Medium 1 150 238.56 17.97 16.93 DPC2 42°51′N 128°05′E 41 中Medium 1 580 379.56 21.55 16.19 DPC3 42°50′N 128°01′E 24 中Medium 1 140 204.71 18.69 15.35 DPC4 42°50′N 128°00′E 22 好Good 1 960 197.59 14.68 12.75 DPC5 42°52′N 128°01′E 37 中Medium 820 248.54 19.31 20.01 JC01 43°19′N 130°34′E 59 差Bad 340 182.71 20.26 26.39 JC02 43°19′N 130°34′E 53 差Bad 430 184.02 19.59 24.12 JC03 43°19′N 130°33′E 58 差Bad 1 820 160.30 12.16 12.20 JC04 43°22′N 130°35′E 56 中Medium 1 480 369.71 16.42 18.47 LD01 43°26′N 130°50′E 53 好Good 810 427.95 20.87 26.04 LD02 43°26′N 130°50′E 57 好Good 1 600 309.28 15.54 16.44 LD03 43°28′N 130°39′E 56 好Good 1 480 172.95 11.71 14.26 LD04 43°28′N 130°40′E 72 中Medium 1 500 231.66 12.32 15.63 LD05 43°31′N 130°41′E 29 中Medium 1 040 285.35 19.33 19.27 LD06 43°31′N 130°41′E 28 差Bad 990 223.46 17.48 18.06 LD07 43°31′N 130°41′E 28 好Good 1 090 223.97 16.98 17.71 LD08 43°31′N 130°41′E 28 差Bad 1 020 248.16 18.30 18.59 注: DPC代表大蒲柴河; JC代表金苍; LD代表六道。下同。Notes: DPC is for Dapuchaihe; JC is for Jincang; LD is for Liudao. The same below. 年轮采样选取了位于林冠层且健康无病害的长白落叶松30~40株,使用生长锥在胸径处(离地面1.3 m)沿东西、南北两个方向各取1个树轮样品。所采样品装入塑料吸管内保存。在实验室内按照标准处理程序进行固定、干燥并用砂纸打磨至年轮界限在显微镜下清晰可见。然后使用LINTAB 6树轮测量系统(Rinntech, Germany)以0.01 mm分辨率精确测量逐年轮宽序列。之后使用COFECHA程序[30]对同一样地中的所有年轮宽度序列进行相关性检验,剔除与主序列相关性差的序列,最后使用ARSTAN程序为剩余序列建立标准年表(表 2)[31]。基于此,得到了所有样地的轮宽指数(Ring width index, RWI)序列用于表征林分的逐年径向生长情况。由于各样地林龄不同,为了分析结果的可比较性,截取了所有样地的RWI序列共有的区间(1986—2015年)用于后续的数据分析。
表 2 样地标准年表统计参数Table 2. Summary statistics for standard chronology of each plot样地
Sample plot共同年区间
Common
interval time
span of year平均敏
感度
Mean
sensitivity标准差
Standard
deviationR1样本间平
均相关系数
Mean
correlations
among all
radiiR2不同树木
之间平均相关系数
Mean
correlations
between treesR3同一树木不同样
本之间平均相关系数
Mean correlations
between trees and
within trees信噪比
Signal-to-noise
ratio总体代表性
Express
population
signal第1特征
向量解释量
PCA1/%DPC1 1990—2016 0.203 0.218 0.447 0.655 0.422 21.195 0.955 0.408 DPC2 1984—2016 0.212 0.188 0.375 0.639 0.368 45.342 0.978 0.384 DPC3 1997—2016 0.215 0.185 0.512 0.550 0.506 21.607 0.956 0.449 DPC4 2000—2016 0.220 0.218 0.369 0.572 0.365 9.884 0.908 0.270 DPC5 1992—2016 0.248 0.260 0.449 0.651 0.446 27.539 0.965 0.388 JC01 1969—2016 0.178 0.203 0.180 0.573 0.172 7.155 0.877 0.511 JC02 1974—2016 0.181 0.208 0.444 0.599 0.440 15.976 0.941 0.487 JC03 1969—2016 0.183 0.184 0.367 0.611 0.363 26.076 0.963 0.548 JC04 1979—2016 0.190 0.199 0.505 0.760 0.501 12.733 0.927 0.288 LD01 1979—2016 0.103 0.157 0.217 0.505 0.213 27.837 0.965 0.395 LD02 1966—2016 0.135 0.281 0.501 0.721 0.494 13.516 0.931 0.429 LD03 1968—2016 0.136 0.161 0.298 0.459 0.295 16.985 0.944 0.419 LD04 1961—2016 0.150 0.159 0.340 0.618 0.337 6.475 0.866 0.527 LD05 1994—2016 0.155 0.149 0.326 0.512 0.322 43.199 0.977 0.491 LD06 1993—2016 0.156 0.201 0.351 0.594 0.343 52.478 0.981 0.520 LD07 1993—2016 0.162 0.328 0.443 0.524 0.435 55.652 0.982 0.531 LD08 1993—2016 0.169 0.166 0.493 0.696 0.490 56.135 0.982 0.528 2.2 历史气候数据
本研究使用了2个来源的气候数据。用于分析树木径向生长与历史气候之间关系所使用的数据来自于东安格利亚大学的CRU TS 3.22高分辨率气候数据集,我们从中提取了1986—2015年间的月均温及帕尔默干旱指数(Palmer drought severity index, PDSI)。其中PDSI是一个基于降水量和温度的反映干燥程度的指标,能够很好反映水分的盈缺。为使分析结果更清晰,按照季节对一年的12个月进行了划分,其中上年12月至当年2月为冬季,3—5月为春季,6—8月为夏季,9—11月为秋季,并将对应月份的温度、PDSI取平均作为季节气候数据。CRU数据的空间分辨率为0.5°(研究区所在纬度,栅格边长约为41 km),由于金苍和六道两个研究地点的距离较近,位于同一栅格内,因此在分析时两地使用的气候数据相同。考虑到树木当年的生长与上年的气候之间存在“滞后效应”[32],因此在数据分析时除了使用当年春季到秋季的气候数据,还使用了上年四季的气候数据。
但上述CRU的气候数据无法反映出不同样地间的气候条件差异。采用Wang等[33]利用国家气象台站数据建立的模型,基于样地经、纬度和海拔估算了各样地的年均温(Annual mean temperature, AMT)、年均降水量(Annual precipitation, AP)。然后使用估算的年均温、降水计算了各样地的潜在蒸散量(Potential evapotranspiration, PET)[33]。研究表明,能量和水分的有效性是影响生产力的关键气候要素[34],这里采用PET和PDSI分别反映能量和水分的有效性。由于无法获取计算PDSI需要的一些数据,因此采用从CRU数据集中提取的PDSI数据(取1986—2015年间PDSI的均值)。
2.3 数据分析
首先分析了研究区近30年的年均温、PDSI的变化趋势,然后对各样地的RWI序列与对应的各季节(上年春季到当年秋季)的平均温度、PDSI序列进行相关性分析。由于季节气候指标较多,因此RWI与季节气候指标的相关系数只能用以初步判断影响年轮—气候关系的因素。为能明确起主要作用的因素,对上述得到的相关系数矩阵进行主成分分析。为研究气候条件和林分因子的相对影响大小,计算了各样地的气候及林分因子对PCA前2轴得分的解释力(R2),同时建立多元线性模型并使用逐步回归筛选得到对年轮—气候关系影响最显著的变量。主成分分析和逐步回归分析均用R软件完成。
3. 结果和分析
3.1 研究区气候的干暖化趋势
本研究中不同地点在空间上有一定距离,也表现在气候上的差异(图 1)。大蒲柴河(DPC)温度较金苍(JC)和六道(LD)林场低,PDSI则较金苍和六道低。即相对金苍和六道,大蒲柴河的气候条件更为干冷,但近30年(1986—2015年)的年气候指标变化仍表现出相似的趋势。两地均以0.014 ℃/a的速率变暖,PDSI也以近似速率下降。结合上述分析的温度和PDSI数据变化趋势,不难发现研究区在近30年呈现明显的干暖化趋势。
![]() 图 1 研究地1986—2015年气候变化趋势Figure 1. Climatic changing trend of research area from 1986 to 2015
图 1 研究地1986—2015年气候变化趋势Figure 1. Climatic changing trend of research area from 1986 to 20153.2 落叶松径向生长与气候变化的关系
建立标准年表,其统计参数如表 2所示,年表的平均敏感度介于0.103~0.248之间,标准差则介于0.149~0.281之间,样本总体代表性高达0.866~0.982,均超过了样本总体代表性(EPS)的阈值0.85[35]。这完全符合吴祥定[27]对于高质量年表的要求,适合进一步用以年轮与气候变化关系的分析。
对标准年表轮宽指数与季节温度、PDSI进行的相关分析表明,落叶松径向生长明显受到气候变化的影响,与上年和当年多个季节的温度和PDSI都有显著的相关性。更为重要的是,不同林龄的林分对气候变化的响应存在明显差异(图 2),温度升高对林龄小的林分有促进生长的作用,而对林龄过大的林分则会产生抑制作用,且林龄越大抑制作用越明显。
在对年轮—气候关系进行主成分分析后,发现PCA 1轴和2轴共解释了63.1%的变异,表明PCA前2轴能较好地反映年轮—气候关系中的主要信息,因此可用于进一步探讨不同样地的气候条件、林分因子等因素的影响。由PCA前2轴载荷系数(表 3)中可以看出,PCA 1的季节PDSI的载荷系数均为负数,且绝对值明显高于温度指标,这说明PCA 1主要反映径向生长对水分历史变化的响应,径向生长受到水分的限制作用,PCA 1得分为负时值越小,受到水分限制作用越显著。而PCA 2的季节温度的载荷系数也多为负数且绝对值明显高于PDSI,主要反映轮宽与温度指标的历史变化的相关性,PCA 2得分的负值越小表示受温度的限制作用越强。
表 3 PCA第1、2轴载荷Table 3. Loading of seasonal climatic factors on PCA 1 and PCA 2季节气候指标
Seasonal climatic factorPCA 1 PCA 2 P_Spr_T — 0.369 P_Aut_T 0.287 -0.358 P_Win_T — -0.472 C_Spr_T 0.272 0.216 C_Sum_T 0.275 — C_Aut _T -0.161 -0.326 P_Aut_T — -0.375 P_Spr_PDSI -0.165 -0.331 P_Sum_PDSI -0.364 — P_Aut_PDSI -0.356 — P_Win_PDSI -0.229 0.241 C_Spr_PDSI -0.382 -0.106 C_Sum_PDSI -0.381 — C_Aut_PDSI -0.305 0.152 注:P表示上年; C表示当年。Spr、Sum、Aut、Win分别表示春、夏、秋、冬季。下同。“—”表示系数绝对值过小。Notes: P means last year; C means current year. Spr, Sum, Aut, Win represent spring, summer, autumn, winter, respectively. The same below. “—” means absolute value of coefficients is too small. 3.3 气候条件、林分因子对年轮—气候关系的影响
从PCA分析结果(图 3)可以看到,由于气候条件的差异,能够区分出各个地点所对应的样地,DPC的样地居于图右侧,而LD的样地则位于左侧,JC的样地多数处于二者之间。结合各个样地的PET数据,PCA 1轴上越往右,对应样地的PET越大,PDSI的箭头也均朝向左边。这表明PCA1轴反映了样地气候条件差异对年轮—气候关系的影响(表 3)。通过计算样地气候条件、林分因子等对PCA轴的单因素解释力(表 4),发现PET和PDSI都与PCA 1得分显著相关,且两者作用方向相反,而林龄、林分密度等林分因子与PCA 1、PCA 2得分的相关性均不显著。
![]() 图 3 轮宽指数与季节气候指标相关系数的主成分分析Figure 3. Principal components analysis of ring width index and seasonal climatic factors表 4 样地气候条件及林分因子对PCA前两轴得分的解释力(R2)Table 4. Explanatory power of climatic factors and stand conditions for the scores of PCA 1 and 2 axis (R2)
图 3 轮宽指数与季节气候指标相关系数的主成分分析Figure 3. Principal components analysis of ring width index and seasonal climatic factors表 4 样地气候条件及林分因子对PCA前两轴得分的解释力(R2)Table 4. Explanatory power of climatic factors and stand conditions for the scores of PCA 1 and 2 axis (R2)变量Variable PCA 1 PCA 2 PDSI Palmer drought severity index -0.503** -0.006 潜在蒸发量Potential evapotranspiration 0.301* -0.024 林龄Stand age -0.065 0.163 林分密度Stand density 0.008 0.004 蓄积量Volume 0.014 0.164 立地条件Stand factor 0.105 0.167 注:*相关性较显著(P<0.05),**表示相关性显著(P<0.01),***表示相关性极显著(P<0.01)。下同。Notes: - means negative correlation. * represents significant correlation at P<0.05 level, ** represents significant correlation at P<0.01 level, *** represents significant correlation at P<0.001 level. The same below. 通过逐步回归筛选相关气候因子和林分因子后的结果(表 5),可以看到PDSI对PCA 1的解释力为50.27%,说明样地的水分条件对年轮—气候关系有重要作用; 对于PCA 2,林龄也显著解释了25.58%的变异,说明林龄也显著影响了径向生长对历史气候变化的响应,进一步支持了图 2的结果。综上结果表明,年轮—气候关系主要受到样地气候条件的影响,其次受到林龄的影响,而蓄积量、林分密度等其余林分因子的影响则相对较弱。
表 5 气候指标及林分条件对PCA前两轴得分的逐步回归分析方差分析Table 5. Stepwise regression analysis of the scores of PCA 1 and PCA 2 with climatic factors and stand conditions项目Item df SS/% Sum Sq Mean Sq F Pr(>F) PCA 1 PDSI Palmerdrought severity index 1 50.27 52.63 52.63 15.57 0.00*** 潜在蒸发量Potential evapotranspiration 1 5.36 5.61 5.61 1.87 0.19 林分密度Stand density 1 7.18 7.51 7.51 2.51 0.14 残差Residuals 13 37.19 38.94 3.00 PCA 2 PDSI Palmerdrought severity index 1 0.56 0.25 0.25 0.12 0.74 林龄Stand age 1 26.48 12.07 12.07 5.67 0.03* 蓄积量Volume 1 12.22 5.57 5.57 2.61 0.13 残差Residual 13 60.74 27.70 2.13 4. 讨论
本研究对处于不同气候和林分条件下的长白落叶松林的年轮—气候关系进行了分析,得到的结果与以往的相关研究存在一些异同。
4.1 气候条件对年轮—气候关系的影响
此前已有长白落叶松的相关研究表明,其径向生长对气候变化的响应与气候密切相关。如于大炮等[13]在长白山北坡的研究发现长白落叶松的年表—气候关系随海拔不同而有显著差异,并与林线位置有关。一般认为,长白落叶松的生长在高海拔主要受气温影响,在低海拔则主要受降水量影响[16, 36]。对兴安落叶松等其他树种的研究,也广泛发现了这种现象,这与低海拔温度较高、蒸散较强,导致的水分有效性较低有关[22, 37]。在本研究中,大 蒲柴河、六道和金苍经纬度差异并不是很大,但PDSI仍然解释了PCA 1高达50.27%的变异(表 5),这充分反映了气候条件确实对年轮—气候关系存在重要影响。本研究中所有样地海拔均不太高(550~800 m),结果表明,PDSI的作用远高于PET,即中低海拔长白山落叶松的年表—气候关系确实主要受水分条件的影响,与上述研究一致。不过,这并不能说明热量不起作用。在表 5中,如果将PET放在PDSI之前进入模型,则PET可显著解释PCA 1的30.05%的变异(表 6)。事实上,所谓气候干暖化常常是由于气候变暖导致的蒸散增加、有效水分减少引起的,未必一定是由于降水量的下降导致[38]。这一点在PCA排序图中也很明显,图 3中各个样地的温度朝PCA 1的右方增大,而PDSI箭头方向则刚好相反,说明PCA 1右侧样地的PDSI较低是由于其较高的温度(蒸散)所导致。综上,本研究中不同地点长白落叶松对历史气候变化响应的差异主要受水分条件的影响,但水分亏缺则随着热量的上升而加剧。这也意味着,随着东北地区今后进一步的气候变暖,长白落叶松(尤其是老龄林)的生长将会受到严重威胁。
表 6 变量顺序不同的方差分析Table 6. Appendix 1 variance analysis of variables in different order项目Item df SS/% Sum Sq Mean Sq F Pr(>F) PCA 1 潜在蒸发量Potential evapotranspiration 1 30.05 31.46 31.46 10.50 0.01* PDSI Palmer drought severity index 1 25.58 26.78 26.78 8.94 0.01* 林分密度Stand density 1 7.18 7.51 7.51 2.51 0.14 残差Residual 13 37.19 38.94 3.00 PCA 2 PDSI Palmer drought severity index 1 0.56 0.25 0.25 0.12 0.74 蓄积量Volume 1 16.34 7.45 7.45 3.50 0.08 林龄Stand density 1 22.36 10.19 10.19 4.78 0.05 残差Residual 13 60.74 27.70 2.13 4.2 林分因子对年轮—气候关系的影响
研究表明,过去30年的气候变暖对不同林龄落叶松的影响有着很大差异(图 2):林龄较小的落叶松,RWI与各季节历史温度变化多呈正相关,说明气候变暖能够促进其生长; 而林龄较大的落叶松则多呈负相关关系,说明气候变暖对其径向生长有明显的负效应。
年轮与历史气候变化的关系除了受气候条件影响之外,还受到林分条件的影响。具体来说,树木径向生长也受竞争以及林分株数密度、树种、林龄等因素的影响[23, 39]。而在本研究中,单从表 3的单因素解释力的结果来看,这些林分因子似乎没有影响。而多元回归模型的结果却表明,在同时考虑其他气候、林分因子时,林龄对年轮—气候关系存在显著影响(表 5)。
一些研究表明,水分对径向生长的限制作用会随林龄增加而加剧,一般认为大树对气候的变化更加敏感[40]。但是,关于林龄的影响机制还存在争议。一方面,随着林龄的增加林分生产力下降是常见的现象,可能说明随着林龄增大、树木活力下降,因此对环境胁迫更加敏感。但随着林龄的增长,还伴随着树木大小(蓄积量)的增加,大树由于蒸腾作用强、对水分和养分的需求大,也可导致其对气候变化更敏感[41]。本研究结果表明,林龄对PCA2轴的解释力最大,解释了26.48%的变异,这和上述研究一致,说明林龄是影响年轮—气候关系的重要因素。表 5中,蓄积量的解释力不显著。但即使是将蓄积量放在林龄之前进入模型(表 6),其解释力在0.05水平上也不显著。因此,本研究结果更多地支持上述前一种解释,即林龄自身对长白落叶松的气候变化响应有着重要作用,而树木大小的作用相对是较弱的。不过,本文的结果与Martínez-Vilalta[41]的结果相反,因此,关于林龄和树木大小的影响,还有待于今后进一步研究才能确定。
还有研究表明,林分密度是影响年轮—气候关系的一个重要因素[42]。而在本研究中,林分密度的解释力较弱且不显著(表 5)。本文中林分密度在340~1 960株/hm2之间,样地间差异足够大,因此可以排除是由于林分密度的梯度不够大导致其解释力不显著。即林分密度对年轮—气候关系的影响相对较弱,这一结果也与以往一些研究不同。因此,林分密度对径向生长的气候变化响应究竟有多大影响,也有待进一步探讨。
4.3 干暖化对长白落叶松林的潜在影响
图 1表明吉林北部山区的确存在明显的干暖化趋势,与以往东北地区研究一致。气候干暖化对东北树木生长的不利影响也得到众多研究证实。比如,对大兴安岭库都尔地区兴安落叶松的研究表明,区域性的干暖化趋势已经影响到当地主要树木的生长[14]。对阔叶红松林的研究也表明,虽然近几十年的气候干暖化对其不同演替阶段的径向生长影响复杂,正负效应都存在,但已经导致红松林生长的明显下降[19]。随着今后干暖化趋势的加剧,可以预料到蒸发、蒸腾作用的加剧从而导致土壤水分有效性的降低。这一威胁对于林龄较大的林分尤为严重,可能导致其严重的生长衰退甚至死亡[38],而成熟林却是天然林保护的重要对象。因此,需要深入研究如何采取适当的措施以减少干暖化的不利影响,以有效保护东北地区的森林生态系统。
-
![]()
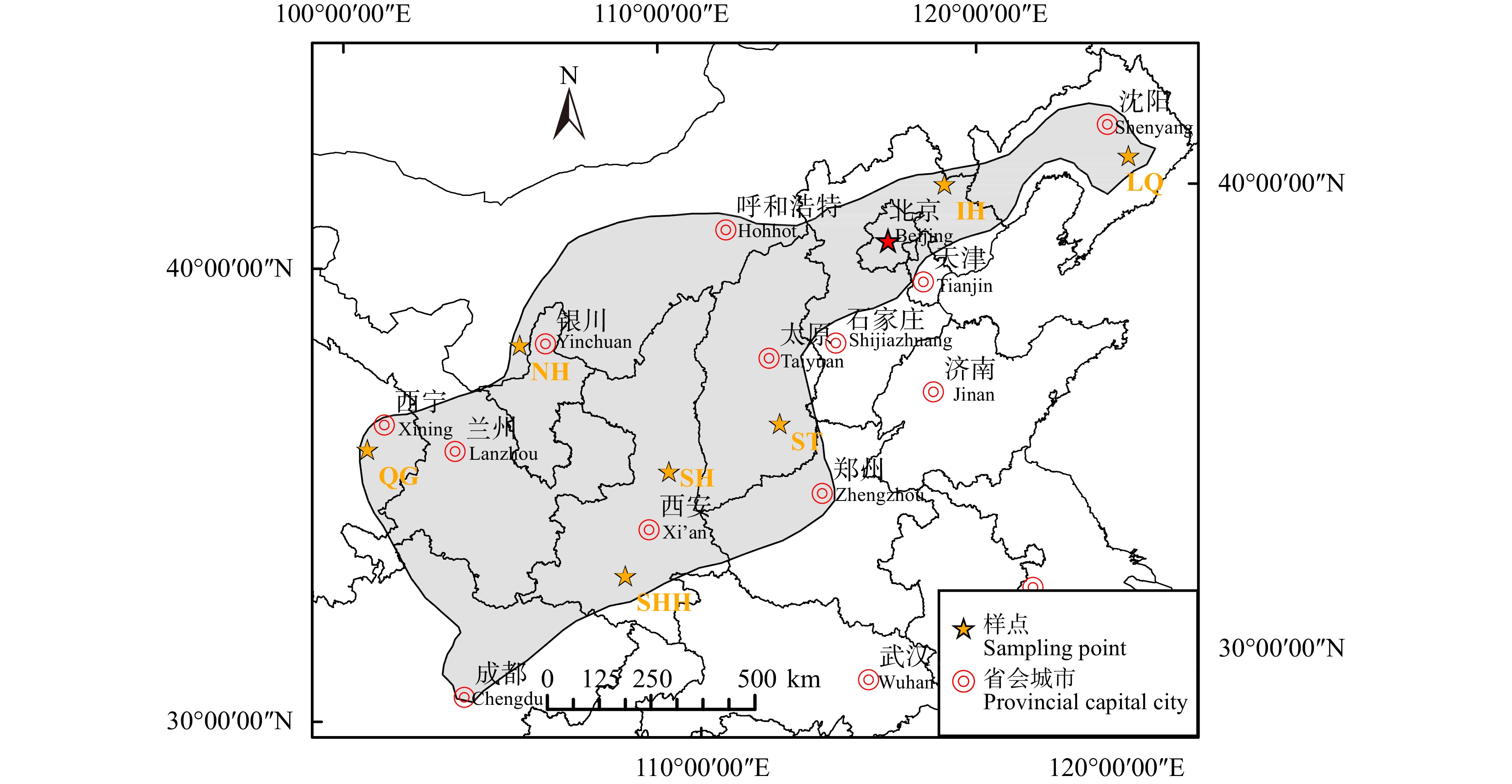
图 1 天然油松林的分布区及样点空间分布
LQ.辽宁千山;IH.内蒙古黑里河;SH.陕西黄龙;ST.山西太岳山;SHH.陕西火地塘;QG.青海贵德;NH.宁夏贺兰山。下同。LQ, Liaoning Qianshan; IH, Inner Mongolia Heili Rever; SH, Shaanxi Huanglong; ST, Shanxi Taiyue Mountains; SHH, Shaanxi Huoditang; QG, Qinghai Guide; NH, Ningxia Helan Mountains. The same below.
Figure 1. Distribution area and spatial distribution of natural P. tabulIformis forests
![]()
图 2 气候和土壤因子对油松叶、枝、根的氮、磷含量影响的方差分解图
A.当年生叶氮;B.当年生叶磷;C.当年生枝氮;D.当年生枝磷;E.多年生叶氮;F.多年生叶磷;G.多年生枝氮;H.多年生枝磷;I.吸收根氮;J.吸收根磷;K.次生根氮;L.次生根磷。a和c分别为气候和土壤的单独解释力,b为两者的协同作用。采用F检验对a、c的显著性进行检验。***表示 P < 0.001;**表示P < 0.01;*表示P < 0.05。下同。A, annual leaf N; B, annual leaf P; C, annual branch N; D, annual branch P; E, perennial leaf N; F, perennial leaf P; G, perennial branch N; H, perennial branch P; I, absorbing root N; J, absorbing root P; K, secondary root N; L, secondary root P. a and c are the pure effects of climate and soil, respectively; b is their synergistic effects. The significances of a and c are evaluated with F test. *** means P < 0.001; ** means P < 0.01; * means P < 0.05. The same below.
Figure 2. Variation partitioning analysis of the effects of climate and soil factors on nitrogen and phosphorus content in leaves, branches, and roots of P. tabuliformis
![]()
图 3 气候和土壤因子对油松叶、枝、根氮磷比的影响的方差分解图
A.当年生叶;B.当年生枝;C.多年生叶;D.多年生枝;E.吸收根;F.次生根。A, annual leaf; B, annual branch; C, perennial leaf ; D, perennial branch; E, absorbing root; F, secondary root.
Figure 3. Variation partitioning analysis of the effects of climate and soil factors on the ratio of nitrogen and phosphorus in leaves, branches, and roots of P. tabuliformis
![]()
图 4 油松叶、枝氮、磷含量随降水的变化
A、E、I.多年生叶;B、F、J.当年生叶; C、G、K.多年生枝;D、H、L.当年生枝。A, E and I, perennial leaves; B, F and J, annual leaves; C, G and K, perennial branches; D, H and L, annual branches.
Figure 4. Changes of nitrogen and phosphorus contents in leaves and branches of P. tabuliformis with precipitation
![]()
图 5 油松叶和枝氮、磷含量随温度的变化
A、E、I.多年生叶;B、F、J.当年生叶; C、G、K.多年生枝;D、H、L.当年生枝。A, E and I, perennial leaves; B, F and J, annual leaves; C, G and K, perennial branches; D, H and L, annual branches.
Figure 5. Changes of nitrogen and phosphorus contents in leaves and branches of P. tabuliformis with temperature
表 1 研究地区自然状况
Table 1 Natural situation of study region
研究地点
Study site经度
Longitude纬度
Latitude海拔
Altitude/m年降水量
Annual
precipitation/
mm年均温度
Annual mean
temperature/℃前一年年
降水量
Annual precipitation
of the previous
year/mm前一年年均温度
Annual mean
temperature of
the previous year/℃土壤氮含量
Soil nitrogen
content/
(mg·g−1)土壤磷含量
Soil phosphorus
content/
(mg·g−1)LQ 123°08′E 40°00′N 260 872 6.45 1 092 6.50 11.05 4.50 IH 118°28′E 41°21′N 1 138 479 7.40 464 7.53 18.98 3.34 SH 109°57′E 35°52′N 1 190 552 10.48 609 11.77 19.84 6.04 ST 112°07′E 36°39′N 1 496 583 9.50 656 10.33 22.98 5.80 SHH 108°28′E 33°26′N 1 643 825 11.09 783 12.45 22.54 5.09 QG 101°36′E 36°05′N 2 804 407 6.40 391 6.72 48.84 2.08 NH 105°55′E 38°44′N 2 131 184 8.81 142 9.82 24.54 4.59  下载: 导出CSV
下载: 导出CSV
表 2 不同地区油松叶、枝、根氮磷化学计量特征比较
Table 2 Comparison of stoichiometric characteristics of nitrogen and phosphorus in leaves, branches and roots of P. tabuliformis from different regions
性状
Trait器官
Organ类型
TypeNH/
(mg·g−1)IH/
(mg·g−1)QG/
(mg·g−1)SH/
(mg·g−1)ST/
(mg·g−1)SHH/
(mg·g−1)LQ/
(mg·g−1)平均
Mean/
(mg·g−1)变异率
Mutation
rate/%F 最小值
Min. value/
(mg·g−1)最大值
Max. value/
(mg·g−1)氮含量
Nitrogen
content叶
Leaf多年生
Perennial13.38 ± 0.94a 11.34 ± 1.45b 11.81 ± 0.89b 10.90 ± 0.94ab 10.72 ± 0.84ab 11.49 ± 1.27b 10.11 ± 0.95a 11.39 ± 1.40 12.29 8.52** 8.14 14.93 当年生
Annual13.06 ± 0.86cd 12.43 ± 1.31bcd 11.16 ± 0.83ab 11.63 ± 1.24ab 12.00 ± 0.98abc 10.71 ± 1.87a 13.53 ± 1.48 12.08 ± 1.53 12.72 5.62** 8.39 15.74 枝
Branch多年生
Perennial7.41 ± 0.82a 5.75 ± 0.73b 5.04 ± 0.88ab 5.78 ± 1.23b 4.54 ± 0.58a 5.72 ± 0.77b 4.81 ± 1.55ab 5.58 ± 1.29 23.17 8.24** 3.21 8.13 当年生
Annual14.98 ± 2.07c 9.17 ± 1.04b 7.06 ± 1.53a 7.63 ± 2.10ab 7.69 ± 1.09ab 7.61 ± 1.02ab 8.92 ± 2.15b 9.01 ± 3.00 33.29 24.82** 5.18 17.94 根
Root吸收根
Absorbing
root8.70 ± 0.74d 5.95 ± 0.44b 5.35 ± 0.93b 4.05 ± 0.79a 6.03 ± 0.53b 7.50 ± 0.99c 8.01 ± 1.21cd 6.51 ± 1.72 26.43 33.44** 2.68 9.93 次生根
Secondary root5.89 ± 0.57b 4.52 ± 0.30a 5.58 ± 0.76b 5.90 ± 1.19b 4.16 ± 0.34a 5.48 ± 0.68b 5.82 ± 1.19b 5.34 ± 1.00 18.76 7.02** 3.55 8.02 磷含量
Phosphorus
content叶
Leaf多年生
Perennial0.98 ± 0.06a 0.73 ± 0.14ab 0.59 ± 0.09a 0.82 ± 0.17b 0.71 ± 0.05ab 0.79 ± 0.23b 0.80 ± 0.10b 0.77 ± 0.172 22.19 6.60** 0.35 1.12 当年生
annual1.46 ± 0.21b 1.44 ± 0.20b 1.00 ± 0.19a 1.03 ± 0.24a 1.45 ± 0.20b 1.39 ± 0.40b 1.21 ± 0.23ab 1.28 ± 0.30 23.85 5.73** 0.66 2.21 枝
Branch多年生
Perennial0.88 ± 0.15d 0.63 ± 0.10ab 0.55 ± 0.14a 0.83 ± 0.15cd 0.62 ± 0.12ab 0.73 ± 0.09bc 0.58 ± 0.09a 0.69 ± 0.17 24.54 9.01** 0.38 1.14 当年生
Annual1.74 ± 0.38c 1.25 ± 0.22b 0.76 ± 0.10a 1.16 ± 0.42b 1.11 ± 0.36b 1.26 ± 0.49b 1.00 ± 0.19ab 1.18 ± 0.42 36.00 7.017** 0.56 2.57 根
Root吸收根
Absorbing
root0.87 ± 0.14d 0.47 ± 0.07bc 0.38 ± 0.06b 0.24 ± 0.07a 0.52 ± 0.05c 0.84 ± 0.18d 0.57 ± 0.20c 0.55 ± 0.24 44.37 29.35** 0.12 1.08 次生根
Secondary root0.52 ± 0.08bc 0.42 ± 0.08b 0.23 ± 0.03a 0.47 ± 0.11b 0.48 ± 0.12b 0.61 ± 0.20c 0.41 ± 0.14b 0.45 ± 0.15 35.23 7.96** 0.19 0.91 氮磷比
N∶P叶
Leaf多年生
Perennial13.64 ± 1.21a 16.17 ± 4.23a 20.49 ± 4.47b 13.81 ± 3.19a 14.98 ± 1.52a 16.03 ± 6.77a 12.77 ± 2.41a 15.41 ± 4.37 28.38 4.01** 9.89 32.67 当年生
Annual9.02 ± 1.10a 8.70 ± 1.29a 11.37 ± 1.56b 11.76 ± 2.93b 8.35 ± 0.98a 8.07 ± 2.05a 11.77 ± 3.57b 9.86 ± 2.56 26.03 5.61** 5.97 18.18 枝
Branch多年生
Perennial8.50 ± 1.24ab 9.17 ± 1.17b 9.30 ± 1.12b 6.93 ± 1.16a 7.45 ± 1.58ab 7.90 ± 1.51ab 8.71 ± 4.03ab 8.28 ± 2.02 24.49 1.85 4.84 15.48 当年生
Annual8.93 ± 2.10bc 7.46 ± 1.42abc 9.17 ± 1.26b 6.98 ± 2.17ab 7.63 ± 2.80abc 6.44 ± 1.41a 8.93 ± 1.46bc 7.93 ± 2.05 25.85 2.94* 3.13 13.46 根
Root吸收根
Absorbing
root10.23 ± 2.07ab 12.65 ± 1.45abc 14.06 ± 1.99bcd 18.20 ± 5.92d 11.63 ± 1.57ab 9.26 ± 2.41a 16.70 ± 9.51cd 13.25 ± 5.29 39.99 4.73** 6.92 36.75 次生根
Secondary root11.50 ± 1.85ab 11.27 ± 2.96ab 23.64 ± 3.46d 12.73 ± 3.06bc 9.02 ± 2.16a 9.79 ± 3.02ab 15.59 ± 6.07c 13.36 ± 5.72 42.80 18.70** 5.85 27.95 注:同行不同小写字母表示不同地区之间差异显著(P < 0.05)。*表示在P < 0.05水平上显著相关;**表示在P < 0.01水平上显著相关。Notes: different lowercase letters in the same line indicate significant differences among different regions (P < 0.05). * means significant correlation at P < 0.05 level; ** means significant correlation at P < 0.01 level.
下载: 导出CSV
表 3 油松根系氮、磷化学计量特征与降水和温度的相关性分析
Table 3 Correlation analysis of nitrogen and phosphorus stoichiometry characteristics of P. tabuliformis root system with precipitation and temperature
气候因子
Climatic factor吸收根氮
Absorbing root N吸收根磷
Absorbing root P次生根氮
Secondary root N次生根磷
Secondary root P吸收根氮比
Absorb root N∶P次生根氮磷比
Absorb root N∶P年降水量
Annual precipitation/mm0.052 −0.008 −0.013 0.166 0.124 −0.120 年均温度
Annual mean temperature/℃−0.086 0.220 −0.006 0.565** −0.184 −0.559** 注:**表示在P < 0.01水平上显著相关。Note: ** means significant correlation at P < 0.01 level.
下载: 导出CSV
-
[1] Yu Q, Chen Q S, Elser J J, et al. Linking stoichiometric homoeostasis with ecosystem structure, functioning and stability[J]. Ecology Letters, 2010, 13(11): 1390−1399. doi: 10.1111/j.1461-0248.2010.01532.x
[2] Song Z L, Liu H Y, Zhao F J, et al. Ecological stoichiometry of N∶P: Si in China’s grasslands[J]. Plant and Soil, 2014, 380(1/2): 165−179.
[3] Roscher C, Thein S, Schmid B, et al. Complementary nitrogen use among potentially dominant species in a biodiversity experiment varies between two years[J]. Journal of Ecology, 2008, 96(3): 477−488. doi: 10.1111/j.1365-2745.2008.01353.x
[4] Han W X, Fang J Y, Guo D L, et al. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytologist, 2005, 168(2): 377−385. doi: 10.1111/j.1469-8137.2005.01530.x
[5] 杨明飞. 我国北方干旱至半湿润区分布的植物叶碳氮磷化学计量特征研究[D]. 兰州: 兰州大学, 2017. Yang M F. Leaf carbon, nitrogen and phosphorus stoichiometry across plant species in the arid to semi-humid regions, North China[D]. Lanzhou: Lanzhou University, 2017.
[6] 赵广帅, 刘珉, 石培礼, 等. 羌塘高原降水梯度植物叶片、根系性状变异和生态适应对策[J]. 生态学报, 2020, 40(1): 295−309. Zhao G S, Liu M, Shi P L, et al. Variation of leaf and root traits and ecological adaptive strategies along a precipitation gradient on Changtang Plateau[J]. Acta Ecologica Sinica, 2020, 40(1): 295−309.
[7] 宗宁, 石培礼, 赵广帅, 等. 降水量变化对藏北高寒草地养分限制的影响[J]. 植物生态学报, 2021, 45(5): 444−455. doi: 10.17521/cjpe.2020.0135 Zong N, Shi P L, Zhao G S, et al. Variations of nitrogen and phosphorus limitation along the environmental gradient in alpine grasslands on the Northern Xizang Plateau[J]. Chinese Journal of Plant Ecology, 2021, 45(5): 444−455. doi: 10.17521/cjpe.2020.0135
[8] Lü X T, Kong D L, Pan Q M, et al. Nitrogen and water availability interact to affect leaf stoichiometry in a semi-arid grassland[J]. Oecologia, 2012, 168(2): 301−310. doi: 10.1007/s00442-011-2097-7
[9] Reich P B, Oleksyn J. Global patterns of plant leaf N and P in relation to temperature and latitude[J]. Proceedings of the National Academy of Sciences, 2004, 101(30): 11001−11006. doi: 10.1073/pnas.0403588101
[10] Tjoelker M G, Reich P B, Oleksyn J. Changes in leaf nitrogen and carbohydrates underlie temperature and CO2 acclimation of dark respiration in five boreal tree species[J]. Plant, Cell & Environment, 1999, 22(7): 767−778.
[11] Han W X, Fang J Y, Reich P B, et al. Biogeography and variability of eleven mineral elements in plant leaves across gradients of climate, soil and plant functional type in China[J]. Ecology Letters, 2011, 14(8): 788−796. doi: 10.1111/j.1461-0248.2011.01641.x
[12] 赵姗宇, 黎锦涛, 孙学凯, 等. 樟子松人工林原产地与不同自然降水梯度引种地土壤和植物叶片生态化学计量特征[J]. 生态学报, 2018, 38(20): 7189−7197. Zhao S Y, Li J T, Sun X K, et al. Responses of soil and plant stoichiometric characteristics along rainfall gradients in Mongolian pine plantations in native and introduced regions[J]. Acta Ecologica Sinica, 2018, 38(20): 7189−7197.
[13] Chen Y H, Han W X, Tang L Y, et al. Leaf nitrogen and phosphorus concentrations of woody plants differ in responses to climate, soil and plant growth form[J]. Ecography, 2013, 36(2): 178−184. doi: 10.1111/j.1600-0587.2011.06833.x
[14] 王凯, 张大鹏, 宋立宁, 等. 氮沉降和降水增加对榆树幼苗不同器官碳氮磷分配格局的影响[J]. 林业科学, 2020, 56(3): 172−183. Wang K, Zang D P, Song L N, et al. Effects of icreasing nitrogen peposition and precipitation on carbon, nitrogen, and phosphorus allocation in different organs of Ulmus pumila seedlings[J]. Scientia Silvae Sinicae, 2020, 56(3): 172−183.
[15] 李俊慧, 彭国全, 杨冬梅. 常绿和落叶阔叶物种当年生小枝茎长度和茎纤细率对展叶效率的影响[J]. 植物生态学报, 2017, 41(6): 650−660. doi: 10.17521/cjpe.2016.0376 Li J H, Peng G Q, Yang D M. Effect of stem length to stem slender ratio of current-year twigs on the leaf display efficiency in evergreen and deciduous broadleaved trees[J]. Chinese Journal of Plant Ecology, 2017, 41(6): 650−660. doi: 10.17521/cjpe.2016.0376
[16] Yao F Y, Chen Y H, Yan Z B, et al. Biogeographic patterns of structural traits and C∶N∶P stoichiometry of tree twigs in China’s forests[J/OL]. PLoS One, 2015, 10(2): e0116391. https://orcid.org/0000-0001-8678-7147.
[17] 邹丽梅. 亚热带6个树种细根形态比较与根序分级构型研究 [D]. 长沙: 中南林业科技大学, 2015. Zou L M. Root identification and varitation in architecture of fine roots of subtropical tree species in southern China[D]. Changsha: Central South University of Forestry and Technology, 2015.
[18] Guo D L, Xia M X, Wei X, et al. Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species[J]. New Phytologist, 2008, 180(3): 673−683. doi: 10.1111/j.1469-8137.2008.02573.x
[19] Freschet G T, Valverde-Barrantes O J, Tucker C M, et al. Climate, soil and plant functional types as drivers of global fine-root trait variation[J]. Journal of Ecology, 2017, 105(5): 1182−1196. doi: 10.1111/1365-2745.12769
[20] Hong J T, Wang X D, Wu J B. Stoichiometry of root and leaf nitrogen and phosphorus in a dry alpine steppe on the Northern Tibetan Plateau[J]. PLoS One, 2018, 9(10): e109052. doi: 10.1371/journal.pone.0109052
[21] 张赟, 赵亚洲, 张春雨, 等. 北京松山油松种群结构及空间分布格局[J]. 应用与环境生物学报, 2009, 15(2): 175−179. Zhang Y, Zhao Y Z, Zhang C Y, et al. Structure and spatial distribution of Pinus tabulaeformis population in the Songshan Nature Reserve, Beijing, China[J]. Chinese Journal of Applied and Environmental Biology, 2009, 15(2): 175−179.
[22] 刘思文, 艾也博, 刘艳红. 北京松山油松叶功能性状沿海拔梯度的变化及其环境解释[J]. 北京林业大学学报, 2021, 43(4): 47−55. Liu S W, Ai Y B, Liu Y H. Variations in leaf functional traits along the altitude gradient of Pinus tabuliformis and its environmental explanations in Beijing Songshan Mountain[J]. Journal of Beijing Forestry University, 2021, 43(4): 47−55.
[23] 田岳梨, 杨航, 王芳玲, 等. 秦岭中段山脊油松叶功能性状差异及其对海拔梯度的响应[J]. 西北植物学报, 2021, 41(2): 300−309. doi: 10.7606/j.issn.1000-4025.2021.02.0300 Tian Y L, Yang H, Wang F L, et al. Difference of leaf functional traits of Pinus tabuliformis and its response to altitude gradient in the middle of Qinling mountains[J]. Acta Botanica Boreali-Occidentalia Sinica, 2021, 41(2): 300−309. doi: 10.7606/j.issn.1000-4025.2021.02.0300
[24] 张凯, 侯继华, 梁冬. 降水梯度对油松天然林内主要植物叶功能性状的影响[J]. 中南林业科技大学学报, 2016, 36(7): 48−54. Zhang K, Hou J H, Liang D. Effect of precipitation gradients on leaf functional traits of main plant in Pinus tabulaeformis forest[J]. Journal of Central South University of Forestry and Technology, 2016, 36(7): 48−54.
[25] 姜沛沛, 曹扬, 陈云明, 等. 不同林龄油松(Pinus tabulaeformis)人工林植物、凋落物与土壤C、N、P化学计量特征[J]. 生态学报, 2016, 36(19): 6188−6197. Jiang P P, Cao Y, Chen Y M, et al. Variation of C, N, and P stoichiometry in plant tissue, litter, and soil during stand development in Pinus tabulaeformis plantation[J]. Acta Ecologica Sinica, 2016, 36(19): 6188−6196.
[26] 黄萍, 刘艳红. 北京松山油松林林分结构和地形对幼苗更新的影响[J]. 生态学杂志, 2018, 37(4): 1003−1009. Huang P, Liu Y H. Effects of stand structure and terrain factors on seedling regeneration of Pinus tabuliformis forest in the Songshan National Nature Reserve, Beijing[J]. Chinese Journal of Ecology, 2018, 37(4): 1003−1009.
[27] 欧晓岚, 刘艳红. 不同坡向及径级油松异龄叶的功能性状[J]. 南京林业大学学报(自然科学版), 2017, 41(4): 80−88. Ou X L, Liu Y H. Effect of age, slope aspects and diameter classes on leaf functional traits of Pinus tabulaeformis in Songshan, Beijing[J]. Journal of Nanjing Forestry University (Natural Science Edition), 2017, 41(4): 80−88.
[28] 张凯. 油松各器官功能性状及其对环境因子响应的研究[D]. 北京: 北京林业大学, 2016. Zhang K. The functional traits of different organs of Pinus tabulaeformis and their response to the environment[D]. Beijing: Beijing Forestry University, 2016.
[29] 鲍士旦. 土壤农化分析[M]. 北京: 中国农业出版社, 2000. Bao S D. Soil agrochemical analysis[M]. Beijing: China Agriculture Press, 2000.
[30] 杨蕾, 孙晗, 樊艳文, 等. 长白山木本植物叶片氮磷含量的海拔梯度格局及影响因子[J]. 植物生态学报, 2017, 41(12): 1228−1238. doi: 10.17521/cjpe.2017.0115 Yang L, Sun H, Fan Y W, et al. Changes in leaf nitrogen and phosphorus stoichiometry of woody plants along an altitudinal gradient in Changbai Mountain, China[J]. Chinese Journal of Plant Ecology, 2017, 41(12): 1228−1238. doi: 10.17521/cjpe.2017.0115
[31] He J S, Wang X P, Flynn D F B, et al. Taxonomic, phylogenetic, and environmental trade-offs between leaf productivity and persistence[J]. Ecology, 2009, 90(10): 2779−2791. doi: 10.1890/08-1126.1
[32] Yuan Z Y, Chen H Y, Reich P B. Global-scale latitudinal patterns of plant fine-root nitrogen and phosphorus[J]. Nature Communications, 2011, 2: 344. doi: 10.1038/ncomms1346
[33] Wang Z Q, Gong H Y, Sardans J, et al. Divergent nitrogen and phosphorus allocation strategies in terrestrial plant leaves and fine roots: a global meta-analysis[J]. Journal of Ecology, 2022, 110: 2745−2758. doi: 10.1111/1365-2745.13985
[34] Reich P B, Tjoelker M G, Pregitzer K S, et al. Scaling of respiration to nitrogen in leaves, stems and roots of higher land plants[J]. Ecology Letters, 2008, 11(8): 793−801. doi: 10.1111/j.1461-0248.2008.01185.x
[35] 周红艳, 吴琴, 陈明月, 等. 鄱阳湖沙山单叶蔓荆不同器官碳、氮、磷化学计量特征[J]. 植物生态学报, 2017, 41(4): 461−470. doi: 10.17521/cjpe.2016.0231 Zhou H Y, Wu Q, Chen M Y, et al. C, N and P stoichiometry in different organs of Vitex rotundifolia in a Poyang Lake desertification hill[J]. Chinese Journal of Plant Ecology, 2017, 41(4): 461−470. doi: 10.17521/cjpe.2016.0231
[36] Pregitzer K S, de Forest J L, Burton A J, et al. Fine root architecture of nine north American trees[J]. Ecological Monographs, 2002, 72(2): 293−309. doi: 10.1890/0012-9615(2002)072[0293:FRAONN]2.0.CO;2
[37] Li A, Guo D L, Wang Z Q, et al. Nitrogen and phosphorus allocation in leaves, twigs, and fine roots across 49 temperate, subtropical and tropical tree species: a hierarchical pattern[J]. Functional Ecology, 2010, 24(1): 224−232. doi: 10.1111/j.1365-2435.2009.01603.x
[38] Bo H J, Wen C Y, Song L J, et al. Fine-root responses of Populus tomentosa forests to stand density[J]. Forests, 2018, 9(9): 562. doi: 10.3390/f9090562
[39] Hishi T. Heterogeneity of individual roots within the fine root architecture: causal links between physiological and ecosystem functions[J]. Journal of Forest Research, 2007, 12(2): 126−133. doi: 10.1007/s10310-006-0260-5
[40] Zadworny M, Eissenstat D M. Contrasting the morphology, anatomy and fungal colonization of new pioneer and fibrous roots[J]. New Phytologist, 2011, 190(1): 213−221. doi: 10.1111/j.1469-8137.2010.03598.x
[41] Williams K, Percival F, Merino J, et al. Estimation of tissue construction cost from heat of combustion and organic nitrogen content[J]. Plant, Cell & Environment, 1987, 10(9): 725−734.
[42] Kerkhoff A J, Fagan W F, Elser J J, et al. Phylogenetic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants[J]. The American Naturalist, 2006, 168(4): E103−E122. doi: 10.1086/507879
[43] Sims D A, Pearcy R W. Response of leaf anatomy and photosynthetic capacity in Alocasia macrorrhiza (Araceae) to a transfer from low to high light[J]. American Journal of Botany, 1992, 79(4): 449−455. doi: 10.1002/j.1537-2197.1992.tb14573.x
[44] Bleecker A B. The evolutionary basis of leaf senescence: method to the madness?[J]. Current Opinion in Plant Biology, 1998, 1(1): 73−78. doi: 10.1016/S1369-5266(98)80131-3
[45] Field C, Mooney H A. Leaf age and seasonal effects on light, water, and nitrogen use efficiency in a California shrub[J]. Oecologia, 1983, 56(2-3): 348−355. doi: 10.1007/BF00379711
[46] 刘万德, 苏建荣, 李帅锋, 等. 云南普洱季风常绿阔叶林优势物种不同生长阶段叶片碳、氮、磷化学计量特征[J]. 植物生态学报, 2015, 39(1): 52−62. doi: 10.17521/cjpe.2015.0006 Liu W D, Su J R, Li S F, et al. Leaf carbon, nitrogen and phosphorus stoichiometry at different growth stages in dominant tree species of a monsoon broad-leaved evergreen forest in Pu’er, Yunnan Province, China[J]. Chinese Journal of Plant ecology, 2015, 39(1): 52−62. doi: 10.17521/cjpe.2015.0006
[47] 汪涛, 杨元合, 马文红. 中国土壤磷库的大小、分布及其影响因素[J]. 北京大学学报(自然科学版), 2008, 44(6): 945−952. Wang T, Yang Y H, Ma W H. The size, distribution and influencing factors of soil phosphorus pools in China storage, patterns and environmental controls of soil phosphorus in China[J]. Acta Scientiarum Naturalium Universitae Pekinensis (Natural Sciences Edition), 2008, 44(6): 945−952.
[48] Smeck N E. Phosphorus dynamics in soils and landscapes[J]. Geoderma, 1985, 36(3−4): 185−199. doi: 10.1016/0016-7061(85)90001-1
[49] 宁志英, 李玉霖, 杨红玲, 等. 科尔沁沙地主要植物细根和叶片碳、氮、磷化学计量特征[J]. 植物生态学报, 2017, 41(10): 1069−1080. doi: 10.17521/cjpe.2017.0048 Ning Z Y, Li Y L, Yang H L, et al. Carbon, nitrogen and phosphorus stoichiometry in leaves and fine roots of dominant plants in Horqin Sandy Land[J]. Chinese Journal of Plant Ecology, 2017, 41(10): 1069−1080. doi: 10.17521/cjpe.2017.0048
[50] 高德新, 张伟, 任成杰, 等. 黄土高原典型植被恢复过程土壤与叶片生态化学计量特征[J]. 生态学报, 2019, 39(10): 3622−3630. Gao D X, Zhang W, Ren C J, et al. Ecological stoichiometry characteristics of soil and leaves during the recovery process of typical vegetation on the Loess Plateau[J]. Acta Ecologica Sinica, 2019, 39(10): 3622−3630.
[51] 姜沛沛, 曹扬, 陈云明, 等. 陕西省3种主要树种叶片、凋落物和土壤N、P化学计量特征[J]. 生态学报, 2017, 37(2): 443−454. Jiang P P, Cao Y, Chen Y M, et al. N and P stoichiometric characteristics of leaves, litter, and soil for three dominant tree species in the Shaanxi Province[J]. Acta Ecologica Sinica, 2017, 37(2): 443−454.
[52] He J S, Fang J Y, Wang Z H, et al. Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grassland biomes of China[J]. Oecologia, 2006, 149(1): 115−122. doi: 10.1007/s00442-006-0425-0
[53] 张慧, 郭卫红, 杨秀清, 等. 麻栎种源林叶片碳、氮、磷化学计量特征的变异[J]. 应用生态学报, 2016, 27(7): 2225−2230. Zhang H, Guo W H, Yang X Q, et al. Variations in leaf C, N, P stoichiometry of Quercus acutissima provenance forests[J]. Chinese Journal of Applied Ecology, 2016, 27(7): 2225−2230.
[54] Dijkstra F A, Blumenthal D, Morgan J A, et al. Contrasting effects of elevated CO2 and warming on nitrogen cycling in a semiarid grassland[J]. New Phytologist, 2010, 187(2): 426−437. doi: 10.1111/j.1469-8137.2010.03293.x
[55] Liu G F, Freschet G T, Pan X, et al. Coordinated variation in leaf and root traits across multiple spatial scales in Chinese semiarid and arid ecosystems[J]. New Phytologist, 2010, 188(2): 543−553. doi: 10.1111/j.1469-8137.2010.03388.x
[56] Mcgroddy M, Daufresne T, Hedin L. Scaling of C∶N∶P stoichiometry in forests worldwide: implications of terrestrial redfield type rations[J]. Ecology, 2004, 85(9): 2390−2401. doi: 10.1890/03-0351
[57] Lovelock C E, Feller I C, Ball M C, et al. Testing the growth rate vs. geochemical hypothesis for latitudinal variation in plant nutrients[J]. Ecology Letters, 2007, 10(12): 1154−1163. doi: 10.1111/j.1461-0248.2007.01112.x
[58] 任书杰, 于贵瑞, 陶波, 等. 中国东部南北样带654种植物叶片氮和磷的化学计量学特征研究[J]. 环境科学, 2007, (12): 2665−2673. doi: 10.3321/j.issn:0250-3301.2007.12.001 Ren S J, Yu G R, Tao B, et al. Leaf nitrogen and phosphorus stoichiometry across 654 terrestrial plant species in NSTEC[J]. Environmental Science, 2007, (12): 2665−2673. doi: 10.3321/j.issn:0250-3301.2007.12.001
-
期刊类型引用(5)
1. 徐金生,姜礅,严善春. Cd胁迫对银中杨幼苗金属元素代谢及其氧化应激的影响. 安徽农业大学学报. 2023(06): 1000-1005 .  百度学术
百度学术
2. 单羽,任晓宁,李雪梅. 非生物胁迫对植物碳水化合物及其代谢相关酶影响的研究进展. 安徽农业科学. 2021(20): 6-9 . 百度学术
3. 熊露露,卢囿佐,刘月炎,彭海兰,黄兴敏,王健健. 外源锌对氮胁迫下薏苡幼苗生长及抗氧化系统的影响. 中国农学通报. 2021(36): 24-29 . 百度学术
4. 刘景华. 不同锌胁迫浓度下杨树生长以及生物量参数的变化. 林业科技通讯. 2020(07): 73-74 . 百度学术
5. 王月月,董效文,姜礅,狄贵秋,王沫,刘晓霞,孟昭军,严善春. 镉、锌复合胁迫对银中杨化学防御的影响. 北京林业大学学报. 2019(06): 96-101 . 本站查看
其他类型引用(5)





计量
- 文章访问数: 489
- HTML全文浏览量: 86
- PDF下载量: 57
- 被引次数: 10
