
Biomass allocation patterns and allometric models of Ginkgo biloba
-
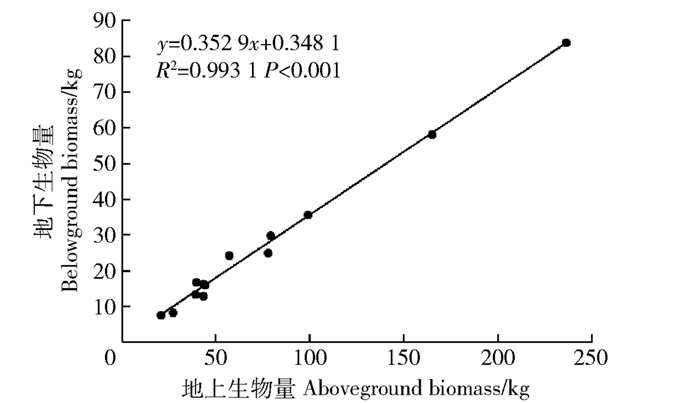
摘要: 以苏北地区银杏人工林为研究对象,选取13株进行整株挖掘,分析不同器官生物量的分配格局,以及地上和地下生物量之间的关系;再分别以胸径(D)、树高(H)、D2H、DaHb为自变量建立银杏各器官生物量模型,选择调整决定系数(Radj2)、残差平方和(SSE)、平均偏差(ME)、平均绝对偏差(MAE)和平均相对误差(MPE)作为选择最优模型的检验指标,根据检验结果筛选出各器官的最优模型。结果表明:13株银杏的整株生物量变化范围为28.50~320.27 kg,树干生物量占总生物量的49.4%~56.6%,树枝生物量占总生物量的12.1%~18.9%,树叶生物量占总生物量的3.8%~5.5%,根生物量占总生物量的26%;地上部分生物量与地下生物量线性方程的斜率为0.35,具有显著的线性相关性(P<0.01);枝和叶生物量都集中于树冠中部,树冠上层和下层的枝、叶生物量明显低于树冠中层生物量(P<0.05),上层和下层生物量之间差异不显著(P>0.05),70%根生物量集中0~1.0 m的土层;枝水平上,基于基径和枝长的枝生物量模型解释量超过95%;在各器官生物量最优模型选择上,以D为自变量的W=aDb的叶、枝、地上部分生物量模型要优于其他模型;树干、根和全株生物量则是以W=aDbHc模型最优。银杏各器官生物量表现为干>根>枝>叶,枝和叶生物量垂直分配上,中冠层占最大比例;基于树高和胸径的相对生长模型可以实现对银杏各器官生物量的准确拟合,银杏生物量及碳储量的有效估算。Abstract: Based on the Ginkgo biloba plantation in northern area of Jiangsu Province, eastern China, 13 sample trees with different diameters at breast height (DBH) were selected, and used to analyze the relationships between above- and below-ground biomass and their allocation patterns. At the individual tree level, allometric models for each component biomass were developed based on independent variables of DBH, tree height (H), D2H and DaHb. The best fitting models were identified by the fitting and test results of parameter estimation, the statistical parameters used in this paper were adjusted determination coefficient (Radj2), sum of squares for error (SSE), statistics estimating the standard deviation SEE, mean relative deviation (ME), mean relative deviation absolute (MAE), mean estimated error (MPE). The results showed that the whole variation range for plant biomass of the 13 ginkgo trees was 28.50-320.27 kg for each tree. Relative proportions of stem, branch, leaf, and root to total tree biomass were 49.4%-56.6%, 12.1%-18.9%, 3.8%-5.5%, and 26%, respectively. The aboveground biomass was significantly linearly correlated with belowground biomass. The slope of the fitted linear model was 0.35. Results showed that the majority leaf and branch biomass occurred in the middle canopy layers, with significant difference between the middle, upper and lower layers in combined biomass of leaves and branches, and there was no significance between upper and lower layers. For all sample trees, about 70% of roots were observed in the 0-1.0 m soil layer. With soil depth increasing, the root biomass decreased exponentially. At branch level, allometric models based on two variables (i.e. BD and BL) of branch biomass explained more than 95% of the variations in data. The results showed that D was a best independent variable in estimating the biomass of leaf, branch, aboveground section than the rest variables, and D-H was the best in estimating stem, root and total tree biomass. The mean value of proportion of different biomass components showed an order of stem > root > branch > leaf. The middle canopy layers occupied the maximum ratio in vertical and horizontal distribution of branch and leaf biomass, and these results were in consistence with the isometric biomass allocation theory. Allometric models based on independent variables of DBH, and H would be suitable for predicting the above- and below-ground component biomass of ginkgo, and the calculation of ginkgo biomass and carbon storage.
-
Keywords:
- biomass /
- allocation pattern /
- allometric model /
- ginkgo plantation
-
-
![]()
图 1 银杏地上生物量与地下生物量之间的关系
Figure 1. Relationship between aboveground and belowground biomass of Ginkgo biloba
![]()
图 2 银杏树干、树枝、树叶、树根生物量空间分布图
Figure 2. Horizontal and vertical distribution of stem, branch, leaf and root biomass of G. biloba
表 1 标准木基本信息表
Table 1 Basic characteristics of sample trees
径级
Diameter at breast height (DBH) class/cm株数
Tree number胸径
DBH/cm树高
Tree height(H)/m树冠长度
Grown length/m南北向冠幅
South-north crown width/m东西向冠幅
East-west crown width/m生物量
Biomass/kg10~15 7 10.9 11.1 6.6 2.6 3.3 28.50 11.4 11.2 9.3 3.2 2.3 35.70 13.3 12.3 10.2 5.3 6.4 56.05 13.4 12.0 9.6 6.0 5.0 56.61 13.5 11.4 7.8 4.9 4.8 52.35 13.6 11.2 8.1 4.9 4.5 59.46 13.8 11.7 9.3 4.7 4.3 60.26 15~20 3 17.1 13.9 11.9 3.9 8.8 81.36 17.7 13.5 9.6 5.1 5.6 102.84 18.8 12.6 10.2 5.7 6.3 108.95 >20 3 20.1 13.0 11.5 6.0 7.2 134.82 24.8 14.5 12.0 6.5 8.6 223.20 27.2 13.6 12.6 8.4 9.7 320.27  下载: 导出CSV
下载: 导出CSV
表 2 株水平上各器官生物量测定值
Table 2 Measurement of each component biomass at the tree level
kg 组分
Component径级DBH class 10~15 cm 15~20 cm >20 cm 叶生物量
Leaf biomass2.62 3.80 17.96 干生物量
Stem biomass28.20 51.59 146.00 枝生物量
Branch biomass6.02 16.01 72.60 地上部分生物量
Aboveground biomass36.84 71.40 236.56 根生物量
Root biomass13.01 26.32 83.70 总生物量
Total biomass49.85 97.72 320.26
下载: 导出CSV
表 3 枝水平上枝生物量异速生长模型
Table 3 Allometric models for branch at branch level
生物量
Biomass/kg模型
Model系数Coefficient Radj2 显著性
Sig.CF SSE MAE MPE a b c 枝Branch lnW=a+blnBD -0.46 ns 3.039*** - 0.904 P<0.001 1.078 1.699 0.279 24.718 lnW=a+blnBD+ClnBL0.039 ns 0.916 ns 2.129 0.957 P<0.001 1.026 0.715 0.201 12.038 注:*表示在0.05水平上差异显著;**表示在0.01水平上差异显著;***表示在0.001水平上差异显著;ns表示在0.05水平上差异不显著;BD表示基径;BL表示枝长。下同。Notes: * means significant difference at P<0.05 level; ** means significant difference at P<0.01 level; *** means significant difference at P<0.01 level; ns means no significant difference at P<0.05 level; BD means branch diameter; BL means branch length. The same below.
下载: 导出CSV
表 4 银杏不同器官生物量模型参数估计、拟合结果和检验结果
Table 4 Parameter estimation, fitting and test results of different components of G. biloba biomass
模型
Model生物量
Biomass/kg系数Coefficient Radj2 显著性Sig. CF SSE MAE MPE a b c lnW=a+blnD 叶Leaf -5.21*** 2.37*** 0.868 P<0.001 1.025 0 0.752 0.199 17.535 干Stem -2.23*** 2.17*** 0.989 P<0.001 1.000 1 0.046 0.048 1.280 枝Branch -6.95*** 3.36*** 0.871 P<0.001 1.077 0 1.471 0.263 26.879 地上Aboveground -2.56*** 2.40*** 0.980 P<0.001 1.003 0 0.104 0.073 1.848 根Root -3.75*** 2.45*** 0.989 P<0.001 1.000 6 0.107 0.078 2.761 全株Total -2.29*** 2.41*** 0.985 P<0.001 1.000 2 0.081 0.066 1.550 lnW=a+blnH 叶Leaf -14.09** 6.13** 0.571 P<0.010 1.075 4 2.518 0.356 33.011 干Stem -10.75** 5.77*** 0.698 P<0.001 1.004 7 1.634 0.297 7.758 枝Branch -19.13** 8.53** 0.550 P<0.001 1.153 1 6.183 0.489 44.667 地上Aboveground -10.51** 5.09*** 0.668 P<0.001 1.005 4 2.166 0.325 7.890 根Root -13.24** 6.46*** 0.678 P<0.001 1.010 8 2.265 0.337 11.212 全株Total -11.55** 6.32*** 0.068 P<0.001 1.004 8 2.159 0.327 7.393 lnW=a+bln(D2H) 叶Leaf -6.95*** 1.03*** 0.853 P<0.001 1.025 4 0.770 0.206 17.974 干Stem -3.84*** 0.95*** 0.980 P<0.001 1.000 2 0.080 0.058 1.556 枝Branch -9.38*** 1.46*** 0.852 P<0.001 1.080 0 1.719 0.288 28.092 地上Aboveground -4.32*** 1.04*** 0.967 P<0.001 1.000 5 0.168 0.082 2.040 根Root -5.60*** 1.07*** 0.967 P<0.001 1.001 0 0.174 0.103 3.542 全株Total -4.07*** 1.05*** 0.972 P<0.001 1.000 3 0.145 0.080 1.823 lnW=a+blnD+clnH 叶Leaf -3.26*** 2.68*** -1.12*** 0.861 P<0.001 1.025 2 0.749 0.200 17.649 干Stem -1.61*** 2.27*** -0.36*** 0.990 P<0.001 1.000 2 0.046 0.480 1.277 枝Branch -3.14*** 3.97*** -2.18*** 0.870 P<0.001 1.077 2 1.377 0.257 26.786 地上Aboveground -1.18*** 2.62*** -0.79*** 0.981 P<0.001 1.000 3 0.100 0.073 1.864 根Root -2.64*** 2.63*** -0.64*** 0.980 P<0.001 1.000 7 0.104 0.074 2.657 全株Total -1.01*** 2.61*** -0.73*** 0.986 P<0.001 1.000 2 0.077 0.067 1.564
下载: 导出CSV
-
[1] OLSON J S, WATTS J A, ALLISON L J. Carbon in live vegetation of major world ecosystems[R]. 1983.
[2] 方精云, 刘国华, 徐嵩龄.我国森林植被的生物量和净生产量[J].生态学报, 1996, 16(5): 497-508. http://www.cqvip.com/Main/Detail.aspx?id=2354606 FANG J Y, LIU G H, XU S L. Biomass and net production of forest vegetation in China[J]. Acta Ecologica Sinica, 1996, 16(5): 497-508. http://www.cqvip.com/Main/Detail.aspx?id=2354606
[3] 国家林业局.第八次全国森林资源清查结果[J].林业资源管理, 2014(1): 1-2. http://d.old.wanfangdata.com.cn/Periodical/lyzygl201401001 State Forestry Administration. The 8th national forestry inventory results[J]. Forest Resources Management, 2014(1): 1-2. http://d.old.wanfangdata.com.cn/Periodical/lyzygl201401001
[4] 冯宗炜.中国森林生态系统的生物量和生产力[M].北京:科学出版社, 1999: 191-196. FENG Z W. Biomass and primary productivity of forest ecosystems in China[M]. Beijing: Science Press, 1999: 191-196.
[5] FANG J Y, WANG Z M. Forest biomass estimation at regional and global levels, with special reference to China's forest biomass[J]. Ecological Research, 2001, 16(3): 587-592. doi: 10.1046/j.1440-1703.2001.00419.x
[6] 罗云建, 张小全, 王效科, 等.森林生物量的估算方法及其研究进展[J].林业科学, 2009, 45(8): 129-134. doi: 10.3321/j.issn:1001-7488.2009.08.023 LUO Y J, ZHANG X Q, WANG X K, et al. Forest biomass estimation methods and their prospects[J]. Scientia Silvae Sinicae, 2009, 45(8): 129-134. doi: 10.3321/j.issn:1001-7488.2009.08.023
[7] 潘维俦, 李利村, 高正衡, 等.杉木人工林生态系统中的生物产量及其生产力的研究[J].湖南林业科技, 1978 (5): 2-14. http://www.cnki.com.cn/Article/CJFD1979-HLKJ197805000.htm PAN W S, LI L C, GAO Z H, et al. Study on biomass and productivity in ecology system of China fir plantation[J]. Hunan Forestry Science and Technology, 1978 (5): 2-14. http://www.cnki.com.cn/Article/CJFD1979-HLKJ197805000.htm
[8] 冯宗炜, 陈楚莹, 张家武, 等.湖南会同地区马尾松林生物量的测定[J].林业科学, 1982, 18(2): 127-134. http://www.cnki.com.cn/Article/CJFDTotal-LYKE198202002.htm FENG Z W, CHEN C Y, ZHANG J W, et al. Determination of biomass of Pinus massoniana stand in Huitong County, Hunan Province[J]. Scientia Silvae Sinicae, 1982, 18(2): 127-134. http://www.cnki.com.cn/Article/CJFDTotal-LYKE198202002.htm
[9] 李文华.小兴安岭谷地云冷杉林群落结构和演替的研究[J].资源科学, 1980, 2(4): 17-29. http://www.cnki.com.cn/Article/CJFDTotal-ZRZY198004002.htm LI W H. Community structure and succession of valley spruce-fir forest in Xiaoxing'an Mountains, China[J]. Natural Resources, 1980, 2(4): 17-29. http://www.cnki.com.cn/Article/CJFDTotal-ZRZY198004002.htm
[10] 李意德, 曾庆波, 吴仲民, 等.尖峰岭热带山地雨林生物量的初步研究[J].植物生态学与地植物学学报, 1992, 16(4): 293-300. http://www.cnki.com.cn/Article/CJFDTotal-ZWSB199204000.htm LI Y D, ZENG Q B, WU Z M, et al. Study on biomass of tropical mountain rain forest in Jianfengling, Hainan Province[J]. Chinese Journal of Plant Ecology, 1992, 16(4): 293-300. http://www.cnki.com.cn/Article/CJFDTotal-ZWSB199204000.htm
[11] 张志, 田昕, 陈尔学, 等.森林地上生物量估测方法研究综述[J].北京林业大学学报, 2011, 33(5): 144-150. http://j.bjfu.edu.cn/article/id/9663 ZHANG Z, TIAN X, CHEN E X, et al. Review of methods on estimating forest aboveground biomass[J]. Journal of Beijing Forestry University, 2011, 33(5): 144-150. http://j.bjfu.edu.cn/article/id/9663
[12] CHUNG S Y, YIM J S, CHO H K, et al. Comparison of forest biomass estimation methods by combining satellite data and field data[J/OL]. Proceedings of IUFRO Division, 2009, 4[2016-08-02]. https://www.researchgate.net/publication/237334259.
[13] 王维枫, 雷渊才, 王雪峰, 等.森林生物量模型综述[J].西北林学院学报, 2008, 23(2): 58-63. http://d.old.wanfangdata.com.cn/Periodical/xblxyxb200802014 WANG W F, LEI Y C, WANG X F, et al. A review of forest biomass models[J]. Journal of Northwest Forestry University, 2008, 23(2): 58-63. http://d.old.wanfangdata.com.cn/Periodical/xblxyxb200802014
[14] 唐守正, 张会儒, 胥辉.相容性生物量模型的建立及其估计方法研究[J].林业科学, 2000, 36(增刊1): 19-27. http://d.old.wanfangdata.com.cn/Periodical/lykx2000Z1003 TANG S Z, ZHANG H R, XU H. Study on establish and estimate method of compatible biomass model[J]. Scientia Silvae Sinicae, 2000, 36(Suppl. 1): 19-27. http://d.old.wanfangdata.com.cn/Periodical/lykx2000Z1003
[15] CHAVE J, ANDALO C, BROWN S, et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests[J]. Oecologia, 2005, 145(1): 712-735. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=687e13a4f79ffd6b30216f5e3963cd74
[16] 曾伟生, 唐守正.一个新的通用性相对生长生物量模型[J].林业科学, 2012, 48(1): 48-52. http://d.old.wanfangdata.com.cn/Periodical/lykx201201009 ZENG W S, TANG S Z. A new general biomass allometric model[J]. Scientia Silvae Sinicae, 2012, 48(1): 48-52. http://d.old.wanfangdata.com.cn/Periodical/lykx201201009
[17] 罗天祥.中国主要森林类型生物生产力格局及其数学模型[D].北京: 中国科学院研究生院, 1996. http://cdmd.cnki.com.cn/Article/CDMD-80061-2006113572.htm LUO T X. Patterns of net primary productivity for Chinese major forest types and their mathematical models[D]. Beijing: Graduate University of the Chinese Academy of Sciences, 1996. http://cdmd.cnki.com.cn/Article/CDMD-80061-2006113572.htm
[18] 左舒翟, 任引, 翁闲, 等.亚热带常绿阔叶林9个常见树种的生物量相对生长模型[J].应用生态学报, 2015, 26(2): 356-362. http://d.old.wanfangdata.com.cn/Periodical/yystxb201502004 ZUO S D, REN Y, WENG X, et al. Biomass allometric equations of nine common tree species in an evergreen broadleaved forest of subtropical China[J]. The Journal of Applied Ecology, 2015, 26(2): 356-362. http://d.old.wanfangdata.com.cn/Periodical/yystxb201502004
[19] MUUKKONEN P. Generalized allometric volume and biomass equations for some tree species in Europe[J]. European Journal of Forest Research, 2007, 126(2): 157-166. doi: 10.1007/s10342-007-0168-4
[20] 曹福亮.中国银杏志[M].北京:中国林业出版社, 2007. CAO F L. Chinese notes of Ginkgo biloba[M]. Beijing: China Forestry Publishing House, 2007.
[21] 陈西娟, 王成章, 叶建中.银杏叶化学成分及其应用研究进展.生物质化学工程[J]. 2008, 42(4): 57-62. doi: 10.3969/j.issn.1673-5854.2008.04.012 CHEN X J, WANG C Z, YE J Z. Research progress of chemical constituents of Ginkgo biloba Linn. leaves and their application[J]. Biomass Chemical Engineering, 2008, 42(4): 57-62. doi: 10.3969/j.issn.1673-5854.2008.04.012
[22] 邵继平, 王伯初, 陈欣, 等.银杏叶提取物药用价值的研究进展[J].重庆大学学报(自然科学版), 2003, 26(1): 130-134. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=cqdxxb200301034 SHAO J P, WANG B C, CHEN X, et al. Advanced research on pharmacology value of the extracts of Ginkgo biloba leaves[J]. Journal of Chongqing University(Natural Science Edition), 2003, 26(1): 130-134. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=cqdxxb200301034
[23] 彭方仁, 张纪林, 李杰, 等.海岸带不同林农复合经营模式的生物生产力研究[J].南京林业大学学报(自然科学版), 2000, 24(2): 78-82. doi: 10.3969/j.issn.1000-2006.2000.02.019 PENG F R, ZHANG J L, LI J, et al. Studies on biomass productivity of three different agroforestry models in seacoast area[J]. Journal of Nanjing Forestry University(Natural Sciences Edition), 2000, 24(2): 78-82. doi: 10.3969/j.issn.1000-2006.2000.02.019
[24] 曹林, 阮宏华, 代劲松, 等.基于HJ-1A/1B CCD数据的区域银杏生物量估测及碳密度制图[J].南京林业大学学报(自然科学版), 2013, 37(2): 8-14. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=njlydxxb201302002 CAO L, RUAN H H, DAI J S, et al. The regional biomass estimation and carbon density mapping of Ginkgo biloba based on HJ-1A/1B CCD satellite image[J]. Journal of Nanjing Forestry University(Natural Sciences Edition), 2013, 37(2): 8-14. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=njlydxxb201302002
[25] 于庚康, 罗艳, 高苹, 等.区域农业经济气象敏感性和气象经济效益[J].生态学杂志, 2012, 31(5): 1265-1271. http://d.old.wanfangdata.com.cn/Periodical/stxzz201205033 YU G K, LUO Y, GAO P, et al. Meteorological sensitivity and meteorological services economic benefit of regional agricultural economy[J]. Chinese Journal of Ecology, 2012, 31(5): 1265-1271. http://d.old.wanfangdata.com.cn/Periodical/stxzz201205033
[26] 孟宪宇.测树学[M].北京:中国林业出版社, 2006: 197-199. MENG X Y. Forest measurement[M]. Beijing: China Forestry Publishing House, 2006: 197-199.
[27] WANG C. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests[J]. Forest Ecology and Management, 2006, 222(1): 9-16. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=d480b7b7505f7c5bcdc3b33c43141c0f
[28] 明安刚, 郑路, 麻静, 等.铁力木人工林生物量与碳储量及其分配特征[J].北京林业大学学报, 2015, 37(2): 32-40. doi: 10.13332/j.cnki.jbfu.2015.02.015 MING A G, ZHEN L, MA J, et al. Biomass and carbon stock and allocation characteristic in Mesua ferrea plantation[J]. Journal of Beijing Forestry University, 2015, 37(2): 32-40. doi: 10.13332/j.cnki.jbfu.2015.02.015
[29] GOWER S T, KUCHARIK C J, NORMAN J M. Direct and indirect estimation of leaf area index, APAR, and net primary production of terrestrial ecosystems[J]. Remote Sensing of Environment, 1999, 70(1): 29-51. doi: 10.1016/S0034-4257(99)00056-5
[30] HALL R J, CASE B S. Erratum: assessing prediction errors of generalized tree biomass and volume equations for the boreal forest region of west-central Canada[J]. Canadian Journal of Forest Research, 2008, 38(6): 878-889. doi: 10.1139/X08-906#.XXMLYPk6s7M
[31] SALIS S M, ASSIS M A, MATTOS P P. Estimating the aboveground biomass and wood volume of savanna woodlands in Brazil's Pantanal Wetlands based on allometric correlations[J]. Forest Ecology & Management, 2006, 228(1): 61-68. https://www.sciencedirect.com/science/article/pii/S0378112706001344
[32] TER-MIKAELIAN M T, KORZUKHIN M D. Biomass equations for sixty-five North American tree species[J]. Forest Ecology & Management, 1997, 97(1): 1-24. doi: 10.1016-S0378-1127(97)00019-4/
[33] ZIANIS D. Predicting mean aboveground forest biomass and its associated variance[J]. Forest Ecology & Management, 2008, 256(6): 1400-1407. http://d.old.wanfangdata.com.cn/NSTLQK/NSTL_QKJJ029672022/
[34] BEETS P N, KIMBERLEY M O, OLIVER G R, et al. Allometric equations for estimating carbon stocks in natural forest in New Zealand[J]. Forests, 2012, 3(3): 818-839. doi: 10.3390/f3030818
[35] NÁVAR J. Allometric equations for tree species and carbon stocks for forests of northwestern Mexico[J]. Forest Ecology & Management, 2009, 257(2): 427-434. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=fe7cbaf2e457a8d05402279feb1f2f4c
[36] WANG C. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests[J]. Forest Ecology & Management, 2006, 222(1): 9-16. https://www.sciencedirect.com/science/article/pii/S0378112705005888
[37] PARRESOL B R. Assessing tree and stand biomass: a review with examples and critical comparisons[J]. Forest Science, 1999, 45(4): 573-593. http://europepmc.org/abstract/AGR/IND22056385
[38] ZABEK L M, PRESCOTT C E. Biomass equations and carbon content of aboveground leafless biomass of hybrid poplar in Coastal British Columbia[J]. Forest Ecology & Management, 2006, 223(1-3): 291-302. https://www.sciencedirect.com/science/article/pii/S0378112705007206
[39] SILESHI G W. A critical review of forest biomass estimation models, common mistakes and corrective measures[J]. Forest Ecology and Management, 2014, 329: 237-254. doi: 10.1016/j.foreco.2014.06.026
[40] NELSON B W, MESQUITA R, PEREIRA J L G, et al. Allometric regressions for improved estimate of secondary forest biomass in the central Amazon[J]. Forest Ecology & Management, 1999, 117(1-3): 149-167. doi: 10.1016-S0378-1127(98)00475-7/
[41] KOZAK A, KOZAK R. Does cross validation provide additional information in the evaluation of regression models?[J]. Canadian Journal of Forest Research, 2003, 33(6): 1499.
[42] PEICHL M, ARAIN M A. Allometry and partitioning of above-and belowground tree biomass in an age-sequence of white pine forests[J]. Forest Ecology & Management, 2007, 253(1-3): 68-80. doi: 10.1016-j.foreco.2007.07.003/
[43] 董点, 林天喜, 唐景毅, 等.紫椴生物量分配格局及异速生长方程[J].北京林业大学学报, 2014, 36(4): 54-63. doi: 10.13332/j.cnki.jbfu.2014.04.013 DONG D, LIN T X, TANG J Y, et al. Biomass allocation patterns and allometric models of Tilia amurensis[J]. Journal of Beijing Forestry University, 2014, 36(4): 54-63. doi: 10.13332/j.cnki.jbfu.2014.04.013
[44] HOUGHTON R A, LAWRENCE K T, HACKLER J L, et al. The spatial distribution of forest biomass in the Brazilian Amazon: a comparison of estimates[J]. Global Change Biology, 2001, 7(7): 731-746. doi: 10.1046/j.1365-2486.2001.00426.x
[45] 汪金松, 张春雨, 范秀华, 等.臭冷杉生物量分配格局及异速生长模型[J].生态学报, 2011, 31(14): 3918-3927. http://d.old.wanfangdata.com.cn/Periodical/stxb201114007 WANG J S, ZHANG C Y, FAN X H, et al. Biomass allocation patterns and allometric models of Abies nephrolepis Maxim.[J]. Acta Ecologica Sinica, 2011, 31(14): 3918-3927. http://d.old.wanfangdata.com.cn/Periodical/stxb201114007
[46] ZHANG H, SONG T, WANG K, et al. Influences of stand characteristics and environmental factors on forest biomass and root-shoot allocation in southwest China[J]. Ecological Engineering, 2016, 91: 7-15. doi: 10.1016/j.ecoleng.2016.01.040
[47] 黄玫, 季劲钧, 曹明奎, 等.中国区域植被地上与地下生物量模拟[J].生态学报, 2006, 26(12): 4156-4163. doi: 10.3321/j.issn:1000-0933.2006.12.031 HUANG M, JI J J, CAO M K, et al. Modeling study of vegetation shoot and root biomass in China[J]. Acta Ecologica Sinica, 2006, 26(12): 4156-4163. doi: 10.3321/j.issn:1000-0933.2006.12.031
[48] LI Z, KURZ W A, APPS M J, et al. Belowground biomass dynamics in the carbon budget model of the Canadian forest sector: recent improvements and implications for the estimation of NPP and NEP[J]. Canadian Journal of Forest Research, 2011, 33(1): 126-136. doi: 10.1139/x02-165#.XXML6fk6s7M
[49] LEVERENZ J W. Shade-shoot structure, photosynthetic performance in the field, and photosynthetic capacity of evergreen conifers[J]. Tree Physiology, 1996, 16(16): 109-114. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=HighWire000001746605
[50] BRIX H. Effects of thinning and nitrogen fertilization on branch and foliage production in Douglas-fir[J]. Canadian Journal of Forest Research, 1981, 10(11): 502-511. doi: 10.1139-x81-069/
[51] XIAO C W, CEULEMANS R. Allometric relationships for below-and aboveground biomass of young Scots pines[J]. Forest Ecology & Management, 2005, 203(1-3): 177-186. doi: 10.1016-j.foreco.2004.07.062/
[52] LACLAU P. Root biomass and carbon storage of ponderosa pine in a northwest Patagonia plantation[J]. Forest Ecology & Management, 2003, 173(1): 353-360. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=bd23d10b04d27e526ac15a2c3d5a8434
[53] NETER J, KUTNER M H, NACHTSHEIM C J, et al. Applied linear statistical model[J]. Journal of the American Statistical Association, 1986, 81: 19-32. http://d.old.wanfangdata.com.cn/NSTLQK/NSTL_QKJJ026998328/
[54] 李轩然, 刘琪璟, 陈永瑞, 等.千烟洲人工林主要树种地上生物量的估算[J].应用生态学报, 2006, 17(8): 1382-1388. doi: 10.3321/j.issn:1001-9332.2006.08.005 LI X R, LIU Q J, CHEN Y R, et al. Aboveground biomass of three conifers in Qianyanzhou plantation[J]. The Journal of Applied Ecology, 2006, 17(8): 1382-1388. doi: 10.3321/j.issn:1001-9332.2006.08.005
[55] 向玮, 雷相东, 刘刚, 等.近天然落叶松云冷杉林单木枯损模型研究[J].北京林业大学学报, 2008, 30(6): 90-98. doi: 10.3321/j.issn:1000-1522.2008.06.014 XIANG W, LEI X D, LIU G, et al. Individual tree mortality models for semi-natural larch-spruce-fir forests in Jilin Province, northeastern China[J]. Journal of Beijing Forestry University, 2008, 30(6): 90-98. doi: 10.3321/j.issn:1000-1522.2008.06.014
[56] MOWRER H T, FRAYER W E. Variance propagation in growth and yield projections[J]. Canadian Journal of Forest Research, 1986, 16(6): 1196-1200. doi: 10.1139/x86-213
-
期刊类型引用(3)
1. 李辉,林沂,孟祥爽,史振伟,蔡万园. 基于地基激光雷达的栾树分形特征分析. 山东农业大学学报(自然科学版). 2022(03): 475-483 .  百度学术
百度学术
2. 何东健,熊虹婷,芦忠忠,刘建敏. 基于多视角立体视觉的拔节期玉米水分胁迫预测模型. 农业机械学报. 2020(06): 248-257 . 百度学术
3. 郭彩玲,刘刚. 基于颜色取样的苹果树枝干点云数据提取方法. 农业机械学报. 2019(10): 189-196 . 百度学术
其他类型引用(7)

计量
- 文章访问数: 3240
- HTML全文浏览量: 782
- PDF下载量: 113
- 被引次数: 10
