
Effects of NaHCO3 stress on growth, photosynthesis and chlorophyll fluorescence characteristics in Populus davidiana × P. bolleana overexpressed TaLEA
-
摘要: 为了筛选耐苏打盐渍土的转基因杨树优良株系, 以苏打盐渍土的主要成分NaHCO3胁迫处理转TaLEA基因的山新杨各株系与对照(NT),测定不同株系的苗高和地径等生长性状,测定净光合速率(Pn)、胞间CO2浓度(Ci)、气孔导度(Gs)、蒸腾速率(Tr)等光合参数以及最大光化学效率(Fv/Fm)、光化学猝灭系数(qP)、非光化学猝灭系数(NPQ)、实际光化学电子产量(ΦPSII)等叶绿素荧光动力学参数,利用染色法分别比较各株系的超氧离子和过氧化氢的累积情况,进而综合评价不同株系对NaHCO3胁迫的响应。结果表明,NaHCO3胁迫处理前、后各株系的苗高和地径生长量发生明显分化,由处理前的差异不显著,到处理20 d后的显著,转基因株系的苗高、地径普遍较对照高,表现突出的是转基因株系SL2号,其平均苗高和地径分别较对照提高了17.4%、15.7%;对照的盐害指数是转基因株系的3.3倍,并且转基因株系的平均存活率是对照的2.8倍。各转基因株系的光合参数和荧光参数也较对照增强,Pn、Tr在胁迫20 d后转基因株系较对照分别高了237.38%、649.02%,Gs、Ci在胁迫15 d时转基因株系较对照分别高了119.05%、24.56%,其中SL2号转基因株系较对照分别提高150%、25.81%。各转基因株系的Fv/Fm平均为0.68,对照只有0.45,前者的qP和ΦPSII只略高于后者,后者的NPQ却是前者的6倍。转基因株系的超氧离子和过氧化氢累积量均少于对照。在相同的碱胁迫环境下,转基因山新杨,尤其是SL2,能保持较快的生长量、较强的光合能力和较低的ROS水平,初步选择其为耐苏打盐渍土的杨树转基因优良株系。Abstract: In order to find the transgenic poplar which can tolerate soda-saline soil, the lines overexpressed TaLEA gene and the control (NT) were treated with NaHCO3, the main components of soda-saline soil. And to evaluate salt-tolerant properties, we measured some parameters of different lines, including height and ground diameter, net photosynthetic rate(Pn), intercellular CO2 concentration(Ci), stomatal conductance(Gs), transpiration rate(Tr), maximal photochemical efficiency(Fv/Fm), photochemical quenching coefficient (qP), non-photochemical quenching coefficient (NPQ), and the actual photochemical electron yield (ΦPSII). Moreover, the accumulation of superoxide anion and hydrogen peroxide in each line was compared by staining method. The results showed that obvious changes had taken place in height and ground diameter among different lines after NaHCO3 stress. There was no significant difference before treatment, but after treating for 20 days, significant differences happened. Compared with control, all transgenic lines increased in height and ground diameter, especially the SL2 transgenic line, whose average height and ground diameter were increased by 17.4% and 15.7%, respectively. Furthermore, the salt damage index of wild type was 3.3 times of transgenic lines, and the survival rate of transgenic lines was 2.8 times of wild type. The photosynthetic parameters and fluorescence parameters of the transgenic lines after 20 days stress were also higher than wild type, Pn and Tr were increased by 237.38% and 649.02%, respectively compared with the wild type. Compared with control, Gs and Ci of transgenic lines were increased by 119.05% and 24.56% under 15 days stress, and Gs, Ci of SL2 transgenic line were increased by 150% and 25.81%. Besides, the average Fv/Fm of each transgenic line was 0.68, the wild type was 0.45. The qP and PSII of transgenic plants were only slightly higher than that of the wild type, but the NPQ of wild type was 6 times of wild type.The accumulation of superoxide anion and hydrogen peroxide in transgenic lines was less than that in wild type. All in all, transgenic lines, especially SL2, could maintain rapid growth, strong photosynthetic capacity and lower level of ROS under alkali stress environment, so SL2 was initially selected as an excellent transgenic line of poplar because of salinity tolerance.
-
Keywords:
- transgene /
- NaHCO3 stress /
- growth /
- photosynthesis /
- chlorophyll fluorescence /
- TaLEA
-
我国是木材消费大国,东北林区作为我国重要的木材生产基地,在满足我国木材需求方面位于重要地位。但是我国的速生丰产林工程从整体上看,存在着很多缺陷,例如数量缺乏,质量不过关,结构不合理等冲突;虽然主要以种植杉松类[1-2]和杨树(Populus spp.)[3]为主的速生丰产林,基本满足了人造板业、造纸业的需求,但对于建筑、家具、装修等所用木材供给还不足[4]。因此,从现有林木资源中筛选出具有速生、丰产优质的造林树种,利用其优良特性营造速生丰产林,对于缓解木材供应的紧张局面具有重要的现实意义。
水冬瓜赤杨(Alnus sibirica)在东北林区广泛分布,具有生长迅速、树干通直的特点[5],与日本落叶松(Larix kaempferi)、山杨(Populus davidiana)、白桦(Butula platyphylla)等速生树种相比,生长速度不相上下,若在同一生境下其直径和材积生长都要比落叶松更快[6]。同时,水冬瓜赤杨还是优良的工业活性炭和制药材料的原料来源[7],并具有较高的涵养水源、保持水土及固氮等生态功能[8],在固堤护岸、水湿地改造以及混交林营建等方面具有重要的应用价值[5],深入研究其种群生态学特征是今后该树种资源开发利用的重要前提和基础。
生命表是种群统计的核心[9-11],存活曲线分析是生命表分析最常用的方法,种群生命表可直接的反映出出生率、死亡率等重要参数,为种群年龄结构、数量动态统计和发展趋势提供重要信息[12-13]。生命表为展现种群结构现状和揭示更新策略提供一定的参考价值,为摸索种群进化演替历程,分析和预测种群动态提供基础[13],并且可以展现种群在群落中的地位与作用以及种群与其所在环境之间的相互联系[10, 14-16]。本文以种群生态学理论为基础,对长白山水冬瓜赤杨种群数量特征进行初步分析,基于空间替代时间的方法,对长白山水冬瓜赤杨种群数量特征进行初步分析,构造水冬瓜赤杨种群生命表,探讨水冬瓜赤杨的生长特征和对策,以期为水冬瓜赤杨种群增长机制研究提供理论依据。
1. 研究地区概况与研究方法
1.1 研究区概况
长白山林区位于吉林省东南部,气候为温带大陆性气候,全区年均降水量为500 ~ 800 mm,降水量的80 %集中于6—9月。土壤类型为森林暗棕壤土、山地棕色针叶林土、山地生草森林土等。经调查发现样地内植被上层乔木主要有:红松(Pinus koraiensis)、白松(Abies holophylla)、落叶松(Larix olgensis)、紫椴(Tilia amurensis)、水曲柳(Fraxinus mandshurica)、花曲柳(Fraxinus rhynchophylla)、白桦(Betula platyphylla)、旱柳(Salix matsudana)、蒙古栎(Quercus mongolica)、赤杨(Alnus japonica)等;中层灌木主要有:刺五加(Eleutherococcus senticosus)、毛榛(Corylus mandshurica)、东北山梅花(Philadelphus schrenkii)等草本植物;下层草本主要为:兴安一枝黄花(Solidago virgaurea)、二叶舞鹤草(Maianthemum bifolium)等[17-18]。
1.2 野外调查方法
根据水冬瓜赤杨种群生长发育过程和生活环境的代表性,对水冬瓜赤杨种群的外业调查分别于2013、2014年进行。根据水冬瓜赤杨的生长范围确定实际调查样地面积,乔木调查利用相邻格子法将样地划分为5 m×5 m的小样方进行,采用每木检尺的方法,记录乔木的胸径、树高和生长状况(病虫害、倾斜、枯立等)。同时记录样地的立地条件如坡位、坡度、坡向、海拔,并准确记录人为干扰情况。
此次调查的群落主要有水冬瓜赤杨—水曲柳混交林、水冬瓜赤杨—旱柳混交林、水冬瓜赤杨—白桦混交林、水冬瓜赤杨林4个类型,合计调查样地数为43块,因部分样地为人工林或未进行年轮条取样,实际选取样地20块,选择的样地面积范围300~900 m2,平均胸径范围7.85~25.60 cm,种群密度范围250~2 650株/hm2,郁闭度范围0.36~0.93,海拔范围304.6~824.7 m,土壤厚度30~85 cm,确保了各种条件下的水冬瓜赤杨群落得到调查,样地概况详情见表 1。
表 1 不同林分样地水冬瓜赤杨种群概况Table 1. General situation of Alnus sibirica population in different plots编号No. 地点Location 样地面积Sample area/m2 种群平均胸径Mean DBH/cm 种群密度/(株·hm-2)Population density/(pant·ha-1) 郁闭度Shade density 海拔Elevat-ion/m 坡位Slope position 坡度Grad-ient 坡向Aspect 土壤厚度Thickness of soil/cm 群落类型Coenotype 1 古洞河林场 600 25.6 250 0.79 824.7 河岸 0° 无 30 水冬瓜赤杨-水曲柳混交林 Gudonghe Forest Farm Riverside None Alnus sibiritca-Fraxinus mandschurica mingled forest 2 东明林场 900 17.38 311 0.77 598.3 河谷 0° 无 34 水冬瓜赤杨-水曲柳混交林 Dongming Forest Farm Valley None Alnus sibirica-Fraxinus mandschurica mingled forest 3 古洞河林场 600 11.58 1 150 0.93 794.1 道边水湿地 0° 无 85 水冬瓜赤杨-旱柳混交林 Gudonghe Forest Farm Roadside meadow bog None Alnus sibirica-Salix matsudana mingled forest 4 古洞河林场 600 10.41 1 050 0.81 795.2 道边水湿地 0° 无 60 水冬瓜赤杨-旱柳混交林 Gudonghe Forest Farm Roadside meadow bog None Alnus sibirica-Salix matsudana mingled forest 5 三合林场 400 8.13 867 0.75 611.6 河谷 0° 无 60 水冬瓜赤杨-旱柳混交林 Sanhe Forest Farm Valley None Alnus sibirica-Salix matsudana mingled forest 6 三合林场 300 8.6 1 350 0.63 612.9 河谷 0° 无 70 水冬瓜赤杨-旱柳混交林 Sanhe Forest Farm Valley None Alnus sibirica-Salix matsudana mingled forest 7 三合林场 400 12.55 1 025 0.88 614.6 河谷 0° 无 60 水冬瓜赤杨-旱柳混交林 Sanhe Forest Farm Valley None Alnus sibirica-Salix matsudana mingled forest 8 东明林场 400 14.97 925 0.64 549 道边水湿地 0° 无 55 水冬瓜赤杨-白桦混交林 Dongming Forest Farm Roadside meadow bog None Alnus sibirica-Betula platyphylla mingled forest 9 东明林场 450 8.33 377 0.64 598 道边水湿地 0° 无 64 水冬瓜赤杨-白桦混交林 Dongming Forest Farm Roadside meadow bog None Alnus sibirica-Betula platyphylla mingled forest 10 许家沟林场 600 18.24 533 0.73 796.3 道边水湿地 0° 无 85 水冬瓜赤杨-白桦混交林 Xujiagou Forest Farm Roadside meadow bog None Alnus sibirica-Betula platyphylla mingled forest 11 许家沟林场 600 15.78 550 0.74 793 道边水湿地 0° 无 80 水冬瓜赤杨-白桦混交林 Xujiagou Forest Farm Roadside meadow bog None Alnus sibirica-Betula platyphylla mingled forest 12 许家沟林场 600 12.2 867 0.75 815.1 道边水湿地 0° 无 75 水冬瓜赤杨-白桦混交林 Xujiagou Forest Farm Roadside meadow bog None Alnus sibirica-Betula platyphylla mingled forest 13 古洞河林场 600 10.35 1 417 0.82 794.6 道边水湿地 0° 无 30 水冬瓜赤杨林 Gudonghe Forest Farm Roadside meadow bog None Alnus sibirica forest 14 东明林场 900 11.98 1 600 0.67 605.9 河谷 0° 无 30 水冬瓜赤杨林 Dongming Forest Farm Valley None Alnus sibirica forest 15 东明林场 900 13.44 1 088 0.65 596.2 河谷 0° 无 30 水冬瓜赤杨林 Dongming Forest Farm Valley None Alnus sibirica forest 16 东明林场 900 15.58 1 088 0.79 570.2 道边水湿地 0° 无 65 水冬瓜赤杨林 Gudonghe Forest Farm Roadside meadow bog None Alnus sibirica forest 17 东明林场 600 8.04 2 366 0.8 578.9 道边水湿地 0° 无 47 水冬瓜赤杨林 Gudonghe Forest Farm Roadside meadow bog None Alnus sibirica forest 18 三合林场 400 14.06 1 244 0.81 304.6 河谷 0° 无 45 水冬瓜赤杨林 Sanhe Forest Farm Valley None Alnus sibirica forest 19 三合林场 400 9.71 2 475 0.89 563.3 道边水湿地 0° 无 40 水冬瓜赤杨林 Sanhe Forest Farm Roadside meadow bog None Alnus sibirica forest 20 三合林场 450 7.85 2 650 0.92 555.1 下坡 21 ° 东坡 45 水冬瓜赤杨林 Sanhe Forest Farm Down grade East Alnus sibirica forest 1.3 径级划分
考虑到外业工作量与水冬瓜赤杨种群生长规律等实际问题,在数据处理时采用“空间代时间”的方法来说明水冬瓜赤杨种群的发育演替问题[19-20]。径级分布作为衡量群落内林间竞争以及生长稳定的主要指标[21],反映了林分结构的基本特点[22-23]。本文根据水冬瓜毛赤杨的生活史特点,基于空间换时间理论,结合研究区水冬瓜毛赤杨的实际年龄结构特征,按2 cm为一个径级段进行整化重新分级,编制了静态生命表、绘制存活曲线,并对动态变化进行了分析。
1.4 静态生命表编制
静态生命表并不能反应种群生活史的全部过程,仅能说明种群世代更迭过程中的某个特定时间,所以可能会出现死亡率为负的情况[12]。Wratten[25]认为,静态生命表分析出现负值会与数据假设技术造成冲突,所以对水冬瓜赤杨种群的调查数据采用滑技术[23, 25]处理。表中的相关指标参照Wartten[24]和王蒙[26]的统计方法。
本文还根据生命表,利用生存率函数S(t)、累计死亡率函数F(t)、死亡密度函数f(t)和危险率函数λ(t)等4个种群生存分析函数[27-30],进一步分析水冬瓜赤杨种群的动态。
1.5 时间序列预测
本文根据调查所得的各龄级水冬瓜毛赤杨数量,采取时间序列分析的移动平均法[26, 29],对水冬瓜赤杨种群2、5、10年后的数量进行预测,推测种群在未来的发展趋势,模型如下:
Mt=1nt∑k=t−n+1xt 式中:t代表径级,n表示准备进行预测的年限,Mt是第n周期的移动平均,即n年后t龄级的种群个体数;xt表示t径级的存活数。
2. 结果与分析
2.1 水冬瓜赤杨种群静态生命表
利用1.4的方法,采用径级结构代替年龄结构,空间代替时间的方法,将径级从小到大的顺序看作是时间顺序关系,编制了水冬瓜赤杨种群静态生命表,结果见表 2。根据表 2可知,水冬瓜赤杨种群小径级死亡量较大,大径级死亡量较小,即水冬瓜赤杨种群死亡量(dx)随径级的增加而逐渐减小;随着径级的增大,总存活数(Tx)和平均存活数(Lx)呈现递减的趋势。水冬瓜赤杨的生命期望(ex)表现出一定的波动性,首先逐渐下降至第8径级,随后出现小幅度上升至第10径级,10径级之后又开始降低。消失率(Kx)在第1径级到第10径级展现出先上升后下降的变化,峰值出现在第9径级,随后又逐渐上升至第20径级。所得结果说明长白山水冬瓜赤杨种群在发展过程中具有波动性,9~10径级出现的波动说明水冬瓜赤杨种群内自疏和种间它疏作用的加剧。
表 2 长白山林区水冬瓜赤杨种群静态生命表Table 2. Static life table of Alnus sibirica population in Changbai Mountain径级
Diameter classax ax′ lx lnlx dx qx Lx Tx ex Kx 1 99 274 1 000 6.908 78 0.079 961 7 495 7.495 0.082 2 122 253 922 6.826 78 0.085 882 6 534 7.090 0.089 3 195 231 843 6.737 78 0.093 804 5 651 6.704 0.098 4 268 210 765 6.639 78 0.103 725 4 848 6.340 0.108 5 314 188 686 6.531 78 0.114 647 4 122 6.008 0.121 6 179 167 608 6.410 78 0.129 568 3 475 5.719 0.138 7 137 145 529 6.271 78 0.148 490 2 907 5.493 0.161 8 120 124 451 6.111 78 0.174 411 2 417 5.362 0.191 9 220 102 372 5.920 78 0.211 333 2 005 5.387 0.237 10 117 81 294 5.683 27 0.093 280 1 672 5.693 0.098 11 95 73 266 5.585 26 0.096 254 1 392 5.226 0.101 12 67 66 241 5.484 26 0.106 228 1 139 4.727 0.112 13 52 59 215 5.372 26 0.119 203 911 4.229 0.126 14 28 52 190 5.246 26 0.135 177 708 3.731 0.145 15 39 45 164 5.101 26 0.156 151 531 3.233 0.169 16 20 38 139 4.932 26 0.184 126 380 2.737 0.204 17 25 31 113 4.729 26 0.226 100 254 2.242 0.256 18 33 24 88 4.473 26 0.292 75 153 1.750 0.345 19 21 17 62 4.128 26 0.412 49 78 1.265 0.531 20 17 10 36 3.597 26 0.700 24 29 0.800 1.204 21 17 3 11 2.393 — — 5 5 0.500 — 注:ax是x径级的存活数;ax′是x径级经匀滑处理后的存活数; lx是x级标准化的存活数; dx是相邻两径级期间标准化的死亡个体数; qx是各径级的死亡率; Lx指平均的存活个体数(从x径级到x+1径级); Tx是个体总数或总寿命(x径级到超过x径级); ex表示进入x径级个体的生命期望寿命; Kx表示消失率。Notes: ax is the standardized survive number of x-class; lx is the standardized survive number of x-level; dx is the standardized dead number of between the two adjacent age classes; qx is the mortality rate at each age level; Lx is the mean of surviving individuals (from x age to x+1 age); Tx is the total number of individuals or the total life span (x age to more than x age); ex indicates the life expectancy of individual entering into x age; Kx means vanishing rate. 2.2 水冬瓜赤杨种群存活曲线
利用表 2水冬瓜赤杨种群生命表的种群存活数对数值(lnlx)及其对应径级绘制了水冬瓜赤杨存活曲线,并利用Deevey[9, 32-34]所构建的林木Ⅱ、Ⅲ型存活曲线的指数和幂函数模型,分别模拟了水冬瓜赤杨的种群存活曲线,所得模型如下。
Ⅱ型:Nx = 7.9367e-0.036(r=0.8843)
Ⅲ型:Nx = 8.7420x-0.229(r=0.7374)
式中:Nx为x径级时的种群存活数,x为径级。
从图 1可以看出,水冬瓜赤杨种群的存活率与径级表现出显著的负相关,随着径级的变大存活率降低,特别是在第9、18、19、20径级更加明显,其他径级变化则较为平稳一致。由模型拟合结果的r值得出,指数模型的相关系数明显较幂函数大,因此指数模型用来描述水冬瓜赤杨种群更加贴切,即Deevey-Ⅱ型存活曲线用来描述长白山水冬瓜赤杨种群更加合适。
![]() 图 1 长白山林区水冬瓜赤杨种群存活曲线lnlx为种群存活数对数值。Figure 1. Survival curve of Alnus sibitica population in Changbai Mountainlnlx is the logaruthm of the survive number of population.
图 1 长白山林区水冬瓜赤杨种群存活曲线lnlx为种群存活数对数值。Figure 1. Survival curve of Alnus sibitica population in Changbai Mountainlnlx is the logaruthm of the survive number of population.水冬瓜赤杨种群死亡率在各径级下大致相等,但仍存在局部波动,在9径级时死亡率较有升高,成活率快速下降,环境筛选和群落自疏强度较大,在进入第10径级至第17径级后水冬瓜存活数量下降趋势趋于稳定,在第18~20径级由于水冬瓜群落已经到了生理死亡的年龄,所以群落存活数又出现了迅速下降的趋势。这与图 2所表示的水冬瓜赤杨种群的死亡率和消失率曲线的变化一致。
从图 2可看出,水冬瓜赤杨种群的死亡率和消失率曲线变化一致。大体上看种群的死亡率和消失率与径级呈现正相关的关系,均随着径级的变大而增加。但是两条曲线变化均存在微小波动,分别在第9径级和第20径时出现级峰值。分析认为由于群落内部个体生长、个体数上升,密度效应变大,群落内部对生长环境与营养条件的需求日益增加导致种内和种间竞争加剧,出现了第1个峰值,随着水冬瓜赤杨种群进入生理死亡年龄,第1个峰值随之出现,种群内个体迅速死亡。
![]() 图 2 水冬瓜赤杨种群死亡率和消失率曲线qx是各径级的死亡率;kx表示消失率。Figure 2. Mortality rate and vanishong rate curves of Alnus sibirica populationqx is the mortality rate at each age level; kx is the rate of disapperance.
图 2 水冬瓜赤杨种群死亡率和消失率曲线qx是各径级的死亡率;kx表示消失率。Figure 2. Mortality rate and vanishong rate curves of Alnus sibirica populationqx is the mortality rate at each age level; kx is the rate of disapperance.2.3 种群生存函数分析
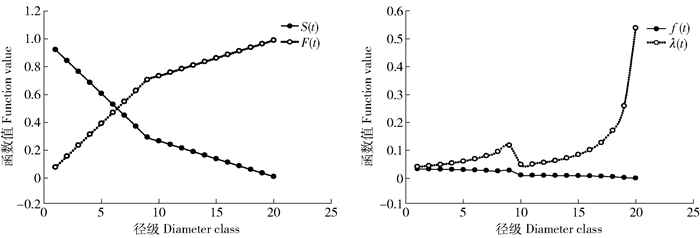
根据水冬瓜赤杨种群静态生命表,分别统计了其种群的生存率函数、累计死亡率函数、死亡密度函数和危险率函数值,绘得图 3。
![]() 图 3 水冬瓜赤杨种群生存函数S(t)为生存率函数;F(t)为累计死亡率函数;f(t)为死亡密度函数;λ(t)为危险率函数。Figure 3. Survival functional rate of Alnus sibirica populationS(t) is the function of available survival rate; F(t) is the cumulative mortality function; f(t) is the death density function; λ(t) is the risk function.
图 3 水冬瓜赤杨种群生存函数S(t)为生存率函数;F(t)为累计死亡率函数;f(t)为死亡密度函数;λ(t)为危险率函数。Figure 3. Survival functional rate of Alnus sibirica populationS(t) is the function of available survival rate; F(t) is the cumulative mortality function; f(t) is the death density function; λ(t) is the risk function.从图 3中可以看出,生存率函数急剧降低,积累死亡率急剧升高,生存函数与积累死亡率曲线变化呈现互补的现象,后期两条曲线均较前期变化幅度缓和,说明水冬瓜赤杨种群生长前期脆弱、死亡率高的特点。从整体上看,死亡密度函数曲线波动性较小,其中,死亡密度在9径级出现较大波动,即水冬瓜赤杨种群在8径级时自疏或它疏作用较强,较弱的个体均被淘汰。危险率函数在1 ~6径级较为平缓,6径级后快速上升呈波动性变化,峰值出现在9、20径级。水冬瓜赤杨种群危险率曲线表现出剧烈的波动性,然而死亡密度曲线变化平缓,且稳中有降。由于大龄的水冬瓜个体数量较少,尽管死亡率进一步上升,但是死亡密度还是逐步降低的,总体来说,危险率均比死亡率高。
综上所述,水冬瓜赤杨的生物学特性和其生活环境共同导致了该种群幼龄时的高死亡率。水冬瓜赤杨种群幼龄时密度可以高达1 000株/ hm2以上,随着个体发育的进行,对光照、水分、空间、营养等环境条件的需求逐渐增长,由于资源有限而竞争压力增强,较弱的个体因其生长要求所需的资源得不到满足而被淘汰,死亡率较高。当水冬瓜成长到其生理死亡年龄,个体对外界干扰的抵抗能力变弱,种群迅速走向消亡。存活曲线、死亡率、消失率和四个生存函数均呈现出前期平稳,中期出现波动,后期衰退的种群特点。
2.4 种群时间序列分析
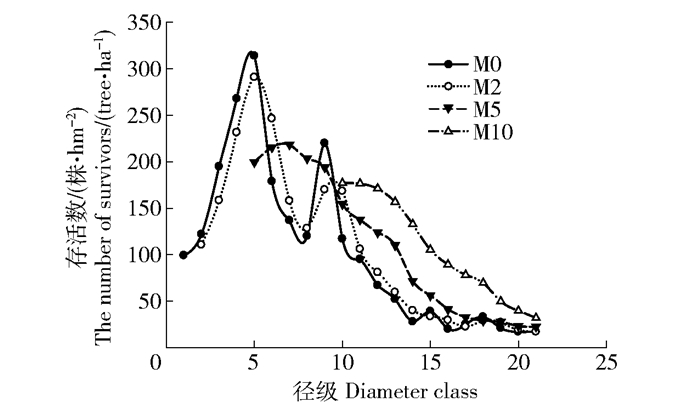
本文以实际调查水冬瓜赤杨种群径级存活数为基础(株/hm2),采用时间序列分析法,并按照一次移动平均法实现[34]来评估预测未来2、5和10年的水冬瓜数量,并绘制时间序列预测图(图 4)。
![]() 图 4 水冬瓜赤杨种群数量动态时间序列预测M0为现在的各径级水冬瓜赤杨数量分布曲线;M2为2年曲线;M5为5年曲线;M10为10年曲线。Figure 4. Time series predictition of Alnus sibitica populationM0 is the curve of current Alnus sibirica quantitative distribution at each age; M2 is the 2-year curve; M5 is the 5-year curve; M10 is the 10-year curve.
图 4 水冬瓜赤杨种群数量动态时间序列预测M0为现在的各径级水冬瓜赤杨数量分布曲线;M2为2年曲线;M5为5年曲线;M10为10年曲线。Figure 4. Time series predictition of Alnus sibitica populationM0 is the curve of current Alnus sibirica quantitative distribution at each age; M2 is the 2-year curve; M5 is the 5-year curve; M10 is the 10-year curve.总整体上看,同一时间曲线的水冬瓜赤杨种群数量均表现出随径级增大而减少的趋势,在未来2、5和10年内,水冬瓜赤杨种群个体数量在幼径级所占比例逐渐下降,在中径级和老径级所占比例逐渐增加,呈明显的衰退趋势。
3. 结论和讨论
3.1 结论
水冬瓜赤杨种群静态生命表的各项指标表明,其种群数量动态存在一定的波动性。种群的存活率随径级的增加逐渐下降,以9径级和20径级的下降较为明显;存活曲线以Deevey-Ⅱ型指数的模拟结果略佳。死亡率和消失率曲线随着径级的增加呈上升趋势,但均存在一定的波动性,峰值分别出现在9径级和20径级。4个生存函数的统计结果表明,随径级的增加种群生存函数急剧降低,积累死亡率急剧升高,死亡密度函数曲线波动性较小,而危险率函数则呈现出较大的波动。综合分析得出,水冬瓜赤杨种群数量表现为前期平稳、中期变化和后期衰退的特征。
同时期的水冬瓜赤杨种群数量曲线均表现出随径级增大而减少的变化趋势,未来2、5和10年的水冬瓜赤杨种群数量幼径级个体数量显著下降,在10年的预测结果中已消失殆尽,而中老径级个体所占比例逐渐增加,种群衰退趋势明显。
3.2 讨论
种群数量动态的波浪式起伏与其生物学特性和环境因素密切相关,是由于个体在生长发育过程中对资源的需求和有限的环境资源之间的矛盾所决定的[35-36]。当种群数量达到了一定的阈值后,随着个体对营养物质和生存空间需求的增加,与邻近个体和上层乔木间的重叠加大,个体间生存空间竞争加剧,环境负载力达到极限,因密度制约使种群产生了自疏和它疏过程。本研究水冬瓜赤杨种群死亡的第1个峰值,即可能归因于种内和种间竞争加剧,而第2个峰值的出现则主要为赤杨种群进入了生理死亡年龄所致,且这种变化在华北落叶松(Larix principis-rupprechtii)[13]和北桑寄生(Coranthus tanakae)种群[37]等研究中也有类似出现。
而水冬瓜赤杨种群衰退趋势明显的原因可能是水冬瓜赤杨为阳性树种,果实的霉烂率高达总量的96.4%,又加上林冠下光照不足,达不到水冬瓜种子发芽所需的最低条件,发芽率低至0.05%[38],造成了种群后备资源不足,缺乏幼径级个体,对其种群演替造成了不利的条件,使其种群衰退。
综上所述,研究得出水冬瓜赤杨种群数量动态存在波动性,种群的天然更新差,表现出明显的衰退特征,属于该地区森林组成的过渡种,今后会被相对稳定的落叶阔叶林群落和地带性顶级群落阔叶红松林所替代。为促进这一自然更替过程,应该根据水冬瓜种群的生长特点,在林下栽种稳定性强的阴性树种,并辅以相应的抚育措施促进群落演替。此外,鉴于本次调查的水冬瓜群落在水湿生境条件下仍能生长良好,并可维持着相对稳定的种群数量性状,可以加以利用进行水湿环境的林地改造,进而为区域的阔叶红松林群落的定居提供良好的环境。
-
![]()
图 1 碱性盐害级值
A.0级无盐害症状;B.1级轻度盐害(有部分叶尖、叶缘变黄);C.2级中度盐害(有大约1/2的叶片、叶缘焦枯);D.3级重度盐害(大部分叶片、叶缘焦枯或脱落);E.4级极重度盐害(复水后植株死亡)。
Figure 1. Level of alkaline salt stress
A, symptom of 0 salt injury; B, level 1 for mild salt damage (the tip of leaf or leaf margin turned yellow); C, level 2 for moderate salt damage (about 1/2 blade or leaf margin turned withered); D, level 3 for severe salt damage (most of the leaf, leaf margin turned withered or was off); E, level 4 for extremely severe salt damage (plant dead after rewatering).
![]()
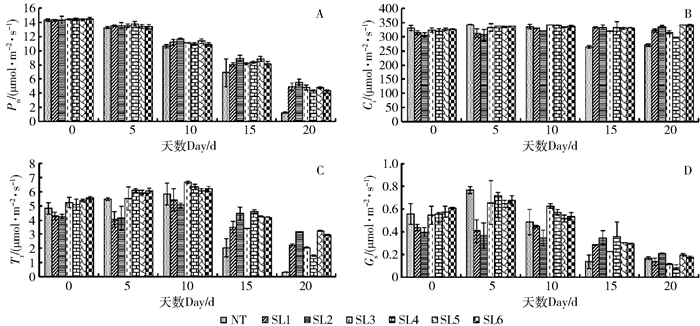
图 2 NaHCO3胁迫对叶片光合参数的影响
A.叶片的净光合速率(Pn);B.胞间CO2浓度(Ci);C.蒸腾速率(Tr);D.气孔导度(Gs)。A, net photosynthetic rate (Pn); B, intercellular CO2 concentration (Ci); C, transpiration rate (Tr); D, stomatal conductance (Gs).
Figure 2. Effects of NaHCO3 stress on photosynthetic parameters of leaves
![]()
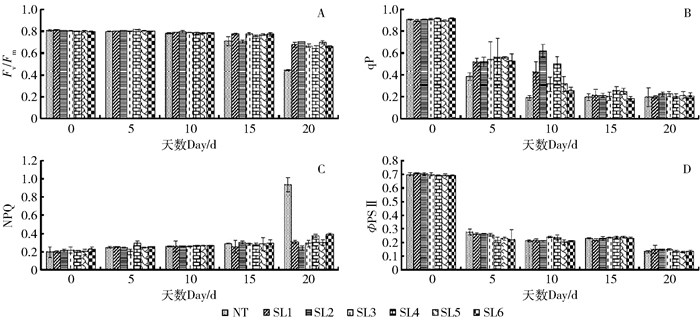
图 3 NaCHO3胁迫对叶片叶绿素荧光动力学参数的影响
A.最大光化学效率(Fv/Fm);B.光化学猝灭系数(qP);C.非光化学猝灭系数(NPQ);D.实际光化学电子产量(ΦPSII)。
Figure 3. Effects of NaCHO3 stress on chlorophyll fluorescence kinetics parameters of leaves
A, maximum photochemical efficiency (Fv/Fm); B, photochemical quenching coefficient (qP); C, non-photochemical quenching coefficient (NPQ); D, actual photochemical electron yield (ΦPSII).
![]()
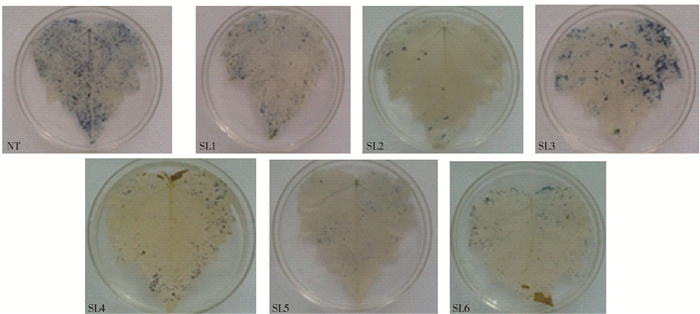
图 4 NaHCO3胁迫下转基因山新杨与NT细胞内超氧离子累积比较
Figure 4. Comparison of superoxide anion accumulation in the cells of transgenic lines and NT under NaHCO3 stress
![]()
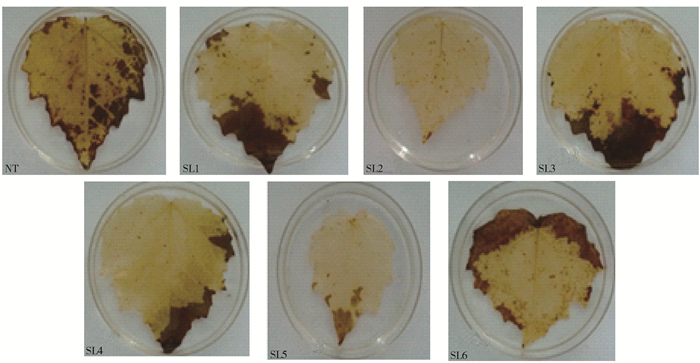
图 5 NaHCO3胁迫下转基因山新杨与NT细胞内过氧化氢累积比较
Figure 5. Comparison of hydrogen peroxide accumulation in transgenic poplar and NT cells under NaHCO3 stress
表 1 NaHCO3胁迫前后高生长比较
Table 1 Comparison in height growth before and after NaHCO3 stress
株系
Line胁迫前、后平均苗高
Average height before and after stress/cm非胁迫条件下
平均苗高
Average height
without stress/cmNaHCO3对高生长的
影响Effect of
NaHCO3 on height
growth/%胁迫前
Before stress胁迫后
After stress绝对高生长
Absolute height growthNT 85.55±3.67 122.11±5.02 c 36.55±5.40 b 147.78±4.42 17.37±2.07ab SL1 84.33±2.36 130.17±2.79 bc 46.00±8.38 ab 147.17±1.01 11.55±1.53ab SL2 88.67±3.21 143.33±7.23 a 54.44±6.16 a 149.67±8.21 4.21±1.75d SL3 85.33±2.60 128.89±6.50 bc 43.56±5.54 ab 145.00±4.58 10.96±7.31bc SL4 85.56±2.14 122.19±2.72 c 37.08±3.83 b 150.67±5.69 18.80±4.36 a SL5 86.30±4.39 132.89±6.44 b 46.92±7.87 ab 139.22±5.42 4.57±1.20 cd SL6 87.05±1.25 127.22±3.53 bc 40.17±2.42 b 143.17±2.56 11.10±3.59ab 平均Mean 86.11±2.80 129.54±4.89 43.53±5.66 146.09±4.55 11.21±2.73 注:不同小写字母表示在P < 0.05水平上差异显著。下同。Notes:different lowercase letters mean significant differences at P < 0.05 level. Same as below.  下载: 导出CSV
下载: 导出CSV
表 2 NaHCO3胁迫前后地径生长比较
Table 2 Comparison in ground diameter growth before and after NaHCO3 stress
株系
Line胁迫前、后平均地径
Average ground diameter before and after stress/mm非胁迫条件下
平均地径Mean
ground diameter
without stress/mmNaHCO3对地径生长
的影响Effect of
NaHCO3 on ground
diameter growth/%胁迫前
Before stress胁迫后
After stress绝对地径生长
Absolute ground diameter growthNT 2.18±0.017 6.48±0.133 c 4.30±0.51 7.53±0.59 13.57±7.71 SL1 2.16±0.10 7.22±0.18 ab 5.06±0.15 8.18±0.63 11.68±8.57 SL2 2.23±0.036 7.50±0.055 a 5.27±0.38 8.49±0.42 11.60±4.02 SL3 2.04±0.058 6.80±0.084 bc 4.77±0.26 7.88±0.63 13.24±7.11 SL4 2.18±0.079 6.92±0.38 bc 4.68±0.35 8.04±0.46 13.87±5.64 SL5 2.18±0.085 6.71±0.36 c 4.53±0.78 7.99±0.47 15.64±8.71 SL6 2.20±0.046 6.79±0.26 bc 4.58±0.35 7.96±0.49 14.50±6.86 平均Mean 2.17±0.063 6.92±0.23 4.74±0.40 8.01±0.52 13.44±6.94
下载: 导出CSV
表 3 碱性盐害指数及存活率
Table 3 Index of salt-alkaline stress and survival rate
株系
Line碱害指数
Index of salt-alkaline stress存活率
Survival rate/%NT 0.79 30 SL1 0.13 87 SL2 0.28 100 SL3 0.30 80 SL4 0.18 90 SL5 0.22 90 SL6 0.34 62
下载: 导出CSV
-
[1] 马驰.松嫩平原土地盐碱化研究[J].吉林农业大学学报, 2014, 36(3):333-337. http://d.old.wanfangdata.com.cn/Periodical/jlnydxxb201403015 MA C. Research on soil salinization in Songnen Plain[J]. Journal of Jilin Agricultural University, 2014, 36(3):333-337. http://d.old.wanfangdata.com.cn/Periodical/jlnydxxb201403015
[2] 张晓光, 黄标, 梁正伟, 等.松嫩平原西部土壤盐碱化特征研究[J].土壤, 2013, 45(2):1332-1338. http://d.old.wanfangdata.com.cn/Periodical/tr201302023 ZHANG X G, HUANG B, LIANG Z W, et al. Study on salinization characteristics of surface soil in western Songnen Plain[J]. Soils, 2013, 45(2):1332-1338. http://d.old.wanfangdata.com.cn/Periodical/tr201302023
[3] 刘正祥, 张华新, 杨秀艳, 等.林木耐盐碱相关基因与基因工程研究进展[J].世界林业研究, 2012, 25(5):11-17. http://d.old.wanfangdata.com.cn/Periodical/sjlyyj201205003 LIU Z X, ZHANG H X, YANG X Y, et al. Research advances in gene and genetic engineering for saline-alkaline tolerance of forest trees[J]. World Forestry Research, 2012, 25(5):11-17. http://d.old.wanfangdata.com.cn/Periodical/sjlyyj201205003
[4] ZHAO X, ZHAN L P, ZOU X Z. Improvement of cold tolerance of the half-high bush Northland blueberry by transformation with the LEA gene from Tamarix androssowii[J]. Plant Growth Regulation, 2011, 63(1):13-22. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=d3893487f7ceac0bb725e5686db22a39
[5] 师静, 刘美芹, 史军娜, 等.沙冬青胚胎晚期发生丰富蛋白基因序列及表达特性分析[J].北京林业大学学报, 2012, 34(4):114-119. http://j.bjfu.edu.cn/article/id/9786 SHI J, LIU M Q, SHI J N, et al. Sequence analysis and expression pattern of AmLEA14 encoding a late embryogenesis abundant protein in Ammopiptanthus mongolicus[J]. Journal of Beijing Forestry University, 2012, 34(4):114-119. http://j.bjfu.edu.cn/article/id/9786
[6] ZHAN J Y, KONG L F, LIU Z P, et al. Stress-induced expression Arabidopsis with a Dehydrin LEA protein from Cleistogenes songorica, a xerophytic desert grass[J]. Plant Omics, 2015, 8(6):485-492.
[7] SUN Y S, HUANG H J, JIANG J, et al. Improved salt tolerance of Populus davidiana×P. bolleana overexpressed LEA from Tamarix androssowii[J]. Journal of Forestry Research, 2014, 25(4):813-818. doi: 10.1007/s11676-014-0529-z
[8] 罗子敬, 孙宇涵, 卢楠, 等.杨树耐盐机制及转基因研究进展[J].核农学报, 2017, 31(3):482-492. http://d.old.wanfangdata.com.cn/Periodical/hnxb201703009 LUO Z J, SUN Y H, LU N, et al. Research advances on salt-tolerance mechanism and genetic transformation of poplar[J]. Journal of Nuclear Agricultural Sciences, 2017, 31(3):482-492. http://d.old.wanfangdata.com.cn/Periodical/hnxb201703009
[9] JAMBUNATHAN N. Determination and detection of reactive oxygen species (ROS), lipid peroxidation, and electrolyte leakage in plants[J]. Methods in Molecular Biology, 2010, 639(1):291-297. http://cn.bing.com/academic/profile?id=f663812894f822f83ae081dfbeaf312b&encoded=0&v=paper_preview&mkt=zh-cn
[10] 倪妍妍, 常二梅, 刘建锋, 等.不同树龄侧柏接穗光合生理的比较研究[J].西北林学院学报, 2017, 32(1):19-24. doi: 10.3969/j.issn.1001-7461.2017.01.03 NI Y Y, CHANG E M, LIU J F, et al. Comparison on photosynthetic physiology in various age scions of Platycladus orientalis[J]. Chinese Academy of Forestry, 2017, 32(1):19-24. doi: 10.3969/j.issn.1001-7461.2017.01.03
[11] 尤鑫, 龚吉蕊.叶绿素荧光动力学参数的意义及实例辨析[J].西部林业科学, 2012, 41(5):90-94. doi: 10.3969/j.issn.1672-8246.2012.05.017 YOU X, GONG J R. Significance and application of chlorophyll fluorescence dynamics process parameters[J]. Journal of West China Forestry Science, 2012, 41(5):90-94. doi: 10.3969/j.issn.1672-8246.2012.05.017
[12] 王智明, 张峰举, 许兴.植物耐盐生理生化指标研究进展[J].湖北农业科学, 2014, 53(7):1493-1496. doi: 10.3969/j.issn.0439-8114.2014.07.003 WANG Z M, ZHANG F J, XU X. Advances on physiological and biochemical indexes of salt tolerance in plant[J]. Hubei Agricultural Sciences, 2014, 53(7):1493-1496. doi: 10.3969/j.issn.0439-8114.2014.07.003
[13] WANG L, ZHANG J L, WANG D, et al. Assessment of salt tolerance in transgenic potato carrying AtNHX1 gene[J].Crop Science, 2013, 53(6):2643-2651. doi: 10.2135/cropsci2013.03.0179
[14] WANG F W, WANG C, SUN Y, et al. Overexpression of vacuolar proton pump ATPase (V-H+ -ATPase) subunits B, C and H confers tolerance to salt and saline-alkali stresses in transgenic alfalfa (Medicago sativa L.)[J]. Journal of Integrative Agriculture, 2016, 15(10):2279-2289. doi: 10.1016/S2095-3119(16)61399-0
[15] 才华, 宋婷婷, 张大洋. rd29A和CaMV-35S启动子调控转AtDREB2A苜蓿耐碱性分析[J].东北农业大学学报, 2016, 47(9):16-23. doi: 10.3969/j.issn.1005-9369.2016.09.003 CAI H, SONG T T, ZHANG D Y. Alkaline tolerance analysis of transgenic alfalfa with AtDREB2A gene regulated by rd29A and CaMV-35S promoter[J]. Journal of Northeast Agricultural University, 2016, 47(9):16-23. doi: 10.3969/j.issn.1005-9369.2016.09.003
[16] DALAL M, TAYAL D, CHINNUSAMY V, et al. Abiotic stress and ABA-inducible group 4 LEA from Brassica napus plays a key role in salt and drought tolerance[J]. Journal of Biotechnology, 2008, 139(2):137-145. http://www.wanfangdata.com.cn/details/detail.do?_type=perio&id=74b85cd7e966d3682afc4169b80ab6cf
[17] YU J, LAI Y, WU X, et al. Overexpression of OsEm1 encoding a group Ⅰ LEA protein confers enhanced drought tolerance in rice[J]. Biochemical and Biophysical Research Communications, 2016, 478(2):703-709. doi: 10.1016/j.bbrc.2016.08.010
[18] 周珩, 郭世荣, 邵慧娟, 等.等渗NaCl和Ca(NO3)2胁迫对黄瓜幼苗生长和生理特性的影响[J].生态学报, 2014, 34(7):1880-1890. http://d.old.wanfangdata.com.cn/Periodical/stxb201407028 ZHOU Y, GUO S R, SHAO H J, et al. Effects of iso-osmotic Ca(NO3)2 and NaCl stress on growth and physiological characteristics of cucumber seedlings[J]. Acta Ecologica Sinica, 2014, 34(7):1880-1890. http://d.old.wanfangdata.com.cn/Periodical/stxb201407028
[19] 卢闯, 逄焕成, 赵长海, 等.水分胁迫下施磷对潮土玉米苗期叶片光合速率、保护酶及植株养分含量的影响[J].中国生态农业学报, 2017, 25(2):239-246. http://d.old.wanfangdata.com.cn/Periodical/stnyyj201702011 LU C, FENG H C, ZHAO C H, et al. Effect of phosphorus on leaf net photosynthesis, protective enzyme activity and nutrient uptake of maize at seedling stage in fluvo-aquic soils under water stress[J]. Chinese Journal of Eco-Agriculture, 2017, 25(2):239-246. http://d.old.wanfangdata.com.cn/Periodical/stnyyj201702011
[20] 许大全.光合作用气孔限制分析中的一些问题[J].植物生理学通讯, 1997, 33(4):241-244. doi: 10.1063-1.1913612/ XU D Q. Some problems in stomatal limitation analysis of photosynthesis[J]. Plant Physiology Communications, 1997, 33(4): 241-244. doi: 10.1063-1.1913612/
[21] MAXWELL K, JOHNSON G. Chlorophyll fluorescence: a practical guide[J]. Journal of Experimental Botany, 2000, 51(345):659-668. doi: 10.1093/jexbot/51.345.659
[22] 寇江涛, 康文娟, 苗阳阳, 等.外源EBR对NaCl胁迫下紫花苜蓿幼苗微量元素吸收及叶绿素荧光动力学参数的影响[J].中国生态农业学报, 2016, 24(3):345-355. http://d.old.wanfangdata.com.cn/Periodical/stnyyj201603009 KOU J T, KANG W J, MIAO Y Y, et al. Effect of exogenous 2, 4-epibrassinolide on trace element absorption and chlorophyll fluorescence of Medicago sativa L. seedlings under NaCl stress[J]. Chinese Journal of Eco-Agriculture, 2016, 24(3):345-355. http://d.old.wanfangdata.com.cn/Periodical/stnyyj201603009
[23] 徐焕文, 刘宇, 姜静, 等.盐胁迫对白桦光合特性及叶绿素荧光参数的影响[J].西南林业大学学报, 2015, 35(4):21-26. http://d.old.wanfangdata.com.cn/Periodical/xnlxyxb201504004 XU H W, LIU Y, JIANG J, et al. Changes of photosynthetic characteristics and chlorophyll fluorescence parameters of Betula platyphylla under salt stress[J]. Journal of Southwest Forestry University, 2015, 35(4):21-26. http://d.old.wanfangdata.com.cn/Periodical/xnlxyxb201504004
[24] 李旭新, 刘炳响, 郭智涛, 等. NaCl胁迫下黄连木叶片光合特性及快速叶绿素荧光诱导动力学曲线的变化[J].应用生态学报, 2013, 24(9):2479-2484. http://d.old.wanfangdata.com.cn/Periodical/yystxb201309012 LI X X, LIU B X, GUO Z T, et al. Effects of NaCl stress on photosynthesis characteristics and fast chlorophyll fluorescence induction dynamics of Pistacia chinensis leaves[J]. Chinese Journal of Applied Ecology, 2013, 24(9):2479-2484. http://d.old.wanfangdata.com.cn/Periodical/yystxb201309012
[25] 张守仁, 高荣孚.光胁迫下杂种杨无性系光合生理生态特性的研究[J].植物生态学报, 2000, 24(5):528-533. doi: 10.3321/j.issn:1005-264X.2000.05.004 ZHANG S R, GAO R F. Ecophysiological characteristics of photosynthesis of hybrid poplar clones under light stress[J]. Acta Phytoecologica Sinica, 2000, 24(5):528-533. doi: 10.3321/j.issn:1005-264X.2000.05.004
[26] 施征, 史胜青, 姚洪军, 等.植物线粒体中活性氧的产生及其抗氧化系统[J].北京林业大学学报, 2009, 31(1):150-154. doi: 10.3321/j.issn:1000-1522.2009.01.026 SHI Z, SHI S Q, YAO H J, et al. Production of ROS and its antioxidant system in plant mitochondria[J]. Journal of Beijing Forestry University, 2009, 31(1):150-154. doi: 10.3321/j.issn:1000-1522.2009.01.026
[27] 戴海芳, 武辉, 阿曼古丽·买买提阿力, 等.不同基因型棉花苗期耐盐性分析及其鉴定指标筛选[J].中国农业科学, 2014, 47(7):1290-1300. doi: 10.3864/j.issn.0578-1752.2014.07.005 DAI H F, WU H, MAIMAITIALI A, et al. Analysis of salt-tolerance and determination of salt-tolerant evaluation indicators in cotton seedlings of different genotypes[J]. Scientia Agricultura Sinica, 2014, 47(7):1290-1300. doi: 10.3864/j.issn.0578-1752.2014.07.005
[28] 李扬, 刘关君, 曲春浦, 等.西伯利亚蓼PsLEA基因的克隆及在NaHCO3胁迫下的表达[J].分子植物育种, 2010, 8(2):276-282. http://d.old.wanfangdata.com.cn/Periodical/fzzwyz201002011 LI Y, LIU G J, QU C P, et al. Cloning of PsLEA gene from Polygonum sibiricum Laxm. and its expression under NaHCO3 stress[J]. Molecular Plant Breeding, 2010, 8(2):276-282. http://d.old.wanfangdata.com.cn/Periodical/fzzwyz201002011
-
期刊类型引用(8)
1. 孙喆,杜玥,王思思. 北方滨海山地公园雨洪管理策略的水文成本效益分析. 风景园林. 2024(06): 99-105 .  百度学术
百度学术
2. 陈泓宇,董宇翔,李雄,刘志成,林辰松. 建筑外环境经济型低影响开发设计模式研究——以南阳一中校园为例. 工业建筑. 2023(04): 54-61 . 百度学术
3. 赖月,康嘉奇,谢怀南,戈晓宇. 半湿润地区公园绿地径流收集灌溉利用成本研究——以迁安市滨湖东路集雨型绿地为例. 北京林业大学学报. 2022(05): 115-123 . 本站查看
4. 周怀宇,刘海龙. 物联网视角下的建成景观设计项目运行信息管理:从空缺到途径. 景观设计学(中英文). 2021(02): 83-95 . 百度学术
5. 禹佳宁,周燕,王雪原,郭诗怡. 城市蓝绿景观格局对雨洪调蓄功能的影响. 风景园林. 2021(09): 63-67 . 百度学术
6. 林俏,刘喆,吕英烁,余曦璇,郑曦. 基于水文模型的北京浅山区雨洪管理措施探究——以夹括河上游为例. 北京林业大学学报. 2020(05): 132-142 . 本站查看
7. 李舟雅,霍锐,戈晓宇. 半湿润地区内源径流型海绵绿地设计方法研究——以山西省晋中市社火公园为例. 中国园林. 2020(06): 107-112 . 百度学术
8. 贾一非,王沛永,田园,迟守冰,王鹏. 高寒地区居住小区海绵化改造建设研究——以西宁市安泰华庭小区为例. 北京林业大学学报. 2019(10): 91-106 . 本站查看
其他类型引用(10)



计量
- 文章访问数: 2037
- HTML全文浏览量: 390
- PDF下载量: 37
- 被引次数: 18
